Research Article - (2013) Volume 2, Issue 2
Dehydrogenases are cellular enzymes usually used as indicators of changes in cell activity and morphology; this includes metabolic processes such as structural differentiation, cell migration, cellular damage and even cell death; hence, assay of enzymes as Lactate dehydrogenase and Glucose 6 phosphate dehydrogenase could provide evidence for the role of some compounds in the stimulation of oxidative stress, cellular damage and eventually cell death.
In this study, examination of Lactate dehydrogenase and Glucose 6 phosphate dehydrogenase in the Prefrontal cortex of Aluminium and Fluoride induced wistar rats was done by administering sodium fluoride (10 mg/kg) and Aluminium Chloride (200 mg/kg). The study employed assay of glucose-6-phospahte dehydrogenase (G6PDH) and lactate dehydrogenase (LDH) levels to determine possible oxidative stress and cellular damage. Results showed alterations in the enzyme levels of G6PDH and LDH indicating a high level of dehydrogenase activity in the treatment groups. This shows that Fluoride and Aluminium have a high potential of causing excitotoxicity, oxidative stress and eventually cellular damage.
Keywords: Prefrontal cortex; Aluminium; Fluoride; Lactate dehydrogenase; Glucose 6 phosphate dehydrogenase
In our everyday life, we are exposed to varying doses of fluoride and aluminium. The former has been shown to be useful in prevention of dental caries (tooth decay), hence it is an important ingredient in toothpaste and is relatively abundant in all forms of water. The latter is sufficiently abundant in household equipments (kitchen utensils), deodorants and used for water treatment. Due to the possible chronic exposure to these substances and their ability to readily interact to form a complex which crosses the blood-brain barrier, it is imperative to assess their neurotoxic effects.
Fluorosis has been described as a condition characterized by the toxicity of elevated fluoride intake on teeth, bones and soft tissues1,2.
The intracellular redox potential plays an important role in cell survival [1]. The principal intracellular reductant NADPH is mainly produced by the pentose phosphate pathway by glucose-6- phosphate dehydrogenase (G6PDH), the rate-limiting enzyme, and by 6-phosphogluconate dehydrogenase [1].
Glucose-6-phosphate dehydrogenase (G6PDH), the rate-limiting enzyme of the pentose phosphate pathway, determines the amount of NADPH by controlling the metabolism of glucose via the PPP [2]. It has been traditionally thought that G6PDH was a typical “housekeeping” enzyme that was regulated solely by the ratio of NADPH to NADP [2]. But research suggests that this enzyme is highly regulated and plays important roles in a variety of cellular processes [3]. This enzyme catalyzes the conversion of glucose-6-phosphate to 6- phosphogluconate, with concomitant production of NADPH and ribose-5-phosphate [4]. The former substance, NADPH, is used as essential substance/substrate for NADPH-dependent ROS producing oxidoreductase enzymes (such as NADPH oxidase and nitric oxide synthase) [1]. G6PD, in one way affecting the redox potential within cells by supplying NADPH itself, and in another way regulating ROS producing enzyme activities, is reported to have a close association with various cell functions/ activities including cell death/necrosis [1]. The latter substance, ribose-5-phosphate, is used in DNA/RNA synthesis within cells [4].
Anaerobic glycolysis and its end product lactate have been synonymous with hypoxic-ischemic hardships ever since the discovery that muscles produce lactate anaerobically. The last step in the glycolytic pathway, the conversion of pyruvate to lactate and the oxidation of NADH to NAD+, is catalyzed by the enzyme lactate dehydrogenase (LDH). LDH is used as a marker of cellular damage [1]. Under aerobic conditions, the NADH would be converted back to NAD+ by the electron transport chain. Since that set of reactions can only happen aerobically, the LDH reaction accomplishes the same end under anaerobic conditions.
Although glucose is usually assumed to be the main energy source for living tissues especially the brain, there are some indications that it is lactate, and not glucose, that is preferentially metabolized by neurons in the brain of several mammals species [5]. According to the lactate-shuttling hypothesis, glial cells are responsible for transforming glucose into lactate, and for providing lactate to the neurons [6]. Because of this local metabolic activity of glial cells, the extracellular fluid immediately surrounding neurons strongly differs in composition from the blood or cerebro-spinal fluid, being much richer with lactate, as it was found in micro dialysis studies [6].
As lactate emerges as an important energy metabolite for brain cells, it follows that the cellular localization of lactate dehydrogenase (LDH, EC 1.1.1.27), the rate-limiting enzyme in lactate fluxes, deserves further attention.
Twenty healthy 5- to 6-week-old adult male albino rats (Wistar), weighing between 100 and 120 g were obtained from the Nigerian Institute of Trypanosomiasis and Onchocerciasis Research, Kaduna, Nigeria and were maintained in the animal house of Bingham University, Karu, Nassarawa State. Approval was done by the ethics committee of the department and animals was treated in accordance to the Guide for Care and Use of Laboratory Animals [7]. They were housed in wooden cages with stainless steel grill tops, and were maintained under proper temperature (25-30ºC), ventilation, and hygienic conditions. They were exposed to 12 hours each of light and dark. The animals were allowed to acclimatize before commencement of the experiment and were maintained on a standard pelleted diet (Vital Feed by GCOML) with water given ad libitum.
Sodium fluoride of 99% purity and aluminium chloride of 97% purity were obtained from SUNLAB Chemicals Ltd, Jos, Plateau State. 1 g of each was dissolved in 50mls of distilled water. The doses administered were prepared from the LD50 values of each compound [8]. LD50 for NaF is 52 mg/kg for rat, while LD50 for AlCl3 is 200-1000 mg/kg in rats [9]. The doses administered were Sodium Fluoride (10 mg/kg body weight) and Aluminium Chloride (200 mg/kg body weight) such that 20% of the LD50 value was used for this study [10].
The animals were divided into four groups of five animals each and weighed before commencement as well as after the experiment. The control group was given tap water [11].
All treatments were given orally with a hypodermic syringe attached to an oral canula. Treatments were administered by 0900(GMT +1) hours daily and the animals were maintained on a standard diet and given water ad libitum throughout the course of the study. The experiment lasted for 21 days and animals were weighed weekly and observed daily. The rats were sacrificed by cervical dislocation on the 22nd day. The brain was excised and placed in 0.25M sucrose solution for biochemical estimation.
The activities of lacate dehydrogenase in the brain increased significantly (p<0.05) as compared with control rats after the treatment with NaF, AlCl3 and their combination (Groups 1-3) for 21 days (Table 1).
| Group no | Treatment | G6PDH | LDH |
|---|---|---|---|
| 1 | NaF | 96.7800 ± 19.35600 | 509.9850 ± 118.726673* |
| 2 | AlCl3 | 164.5260 ± 0.0000 | 1242.5825 ± 319.75250* |
| 3 | NaF + AlCl3 | 179.0430 ± 17.74300* | 3092.2900 ± 828.38833* |
| 4(Control) | Tap water | 169.3650 ± 20.96900 | 279.2775 ± 84.99750 |
Values are mean ± S.E. * represents a significantly (p<0.05) higher mean value
of fluoride in the neural tissue of the fluoride group compared to the control group11
Table 1: G6PDH and LDH activity (u/l) in brain of control and treated rats.
The activities of glucose-6-phosphate dehydrogenase in the brain decreased as compared with control rats after treatment with NaF and AlCl3 (Groups 1 and 2) though, this decrease was not statistically significant. However, there was an increase in the level of G6PD which was statistically significant (p<0.05) after treatment with the combination of NaF and AlCl3 (Group 3) both for 21 days.
Lactate dehydrogenase (LDH) is a cytoplasmatic enzyme present in essentially all major organ systems [11]. Lactate dehydrogenase (LDH, EC 1.1.1.27) is a hydrogen transfer enzyme that catalyses the oxidation of L-lactate to pyruvate with nicotinamide-adenine dinucleotide (NAD+) as hydrogen acceptor, the final step in the metabolic chain of anaerobic glycolysis [12]. The reaction is reversible and the reaction equilibrium strongly favours the reverse reaction, namely the reduction of pyruvate to lactate [13] (Figure 1).
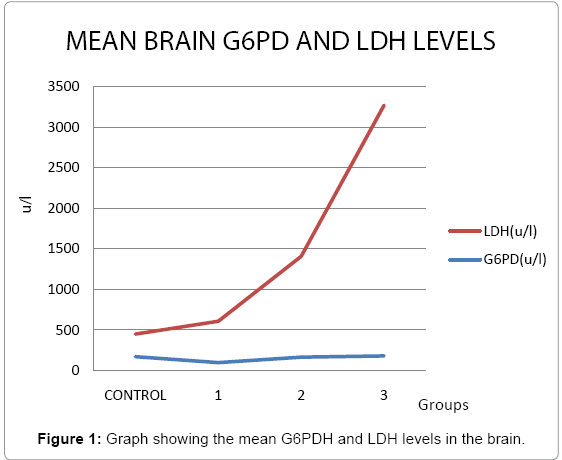
Figure 1: Graph showing the mean G6PDH and LDH levels in the brain.
The extracellular appearance of LDH is used to detect cell damage or cell death [14,15] and its extracellular activity increases under the condition of oxidative stress [12] due to the integrity of cell membrane destroying in the course of lipid peroxidation which allows leakage of the enzyme from even a small mass of damaged tissue [16]. This might explain the increase in LDH levels in all the treatment rats.
G6PDH is a cytoplasmic enzyme that has been used as a direct indicator of oxidative stress [17]; It catalyses the conversion of glucose-6-phosphate (G6P) into 6-phosphogluconate while reducing NADP+ to NADPH in the pentose phosphate pathway [18]. NADPH is physiologically important as it ensures the reduction of glutathione (GSSG to GSH) which is an enzyme in the anti-oxidant defence system [18].
A reduction in G6PDH usually termed as oxidative stress does not imply oxidative stress in its entirety but rather basic changes observed as result of effects of toxicity on the metabolic machinery of the neuron secondary to the oxidative stress [19]. The level of G6PDH are dependent on the composition of the cytoplasm (neuronal metabolic system) such that when oxidative stress is induced, the system will self-regulate to compensate for energy needs i.e. reduction in ATP production will cause G6PDH to withdraw to favour ATP production from pyruvate to meet the energy requirement of the neuron under stress [19]. Hence, a reduction in G6PDH will be observed (as seen in treatment groups 1 and 2) to allow more G6P to proceed to the end of glycolysis to form high energy pyruvate: a precursor of the tricarboxylic acid (TCA) cycle which also explains the lower levels of LDH [19] seen in these treatment groups. However, an increase in G6PDH level as seen in treatment group 3 will imply that more G6P has been shunted to the Pentose Phosphate Pathway to generate ribose sugar for DNA production hence less will proceed to the end of glycolysis which will favour anaerobic glycolysis as seen in the higher level of LDH [19].
The emerging picture from this investigation as well as others conducted on fluoride and aluminium is that there exists a narrow margin between the recommended intakes (1 ppm) and the neurotoxic doses. Therefore, continued monitoring of the exposure of humans to fluoride and aluminium from all sources is required.
We would like to thank Engr Akinrinade Biola (Drakes Technologies, Nigeria) for providing IT support and Dr. Olanrewaju Ajetumobi (Snr Reg Histopathology Dept, UNIJOS, Nigeria) for assisting in microscopic examinations.
