Research Article - (2012) Volume 1, Issue 1
The adult testis is the male reproductive organ that produces sex hormones for sexual behaviours and sperm in a well-controlled process called spermatogenesis. Spermatogenesis involves self-renewal and differentiation of the male germ stem cells spermatogonia, culminating in the production of sperm for germline transmission. Retinoic acid (RA) is essential for fate decision and meiosis entry of mammalian germ cells. The microphthalmia-associated transcription factor (Mitf) is a newly identified transcriptional activator of germ genes in the fish medaka (Oryzias latipes). Here we report an essential role of RA on Mitf-activated germ gene expression and spermatogenesis. Medaka germ gene promoters DAZ, DND and VAS were found to contain RA-responsive elements and to exhibit little Mitf-activated transcriptional activity in response to RA treatment in both medaka ES cells and male germ stem cells. A procedure for testicular organ culture was established, which allows for test-tube spermatogenesis over a 21-day period. By the ordered testicular architecture, cellular morphology and VAS-driven GFP expression, different spermatogenic stages were identifiable on testicular cross-sections. In organ culture, RA treatment generated testes completely free of spermatogonia, and reduced VAS-driven GFP expression in spermatogonia but enhanced its expression in meiotic and post-meiotic germ cells. Therefore, RA represses Mitf-activated germ gene expression in stem cell cultures and differentially regulates germ gene expression depending upon the stages of spermatogenesis, and represses spermatogonial stem cell maintenance in cultured testes of medaka as a lower vertebrate model. These results demonstrate that RA plays a highly conserved role in germ cell development from fish to mammals.
Keywords: Gene expression; Fertility; Retinoids; Organ culture; Spermatogenesis
mitf- Microphthalmia-Associated Transcription Factor; dpt- Day(S) Post Transfection
In diverse animal species, adult gonads, the testis in male and ovary in female, are the reproductive organ that produces sexual hormone for secondary sex characteristics and gametes, sperm in male and eggs in female, for germline transmission. Germline development commences with specification of primordial germ cells (PGCs) early in embryonic development and culminates in the production of gametes in the adult gonads. PGCs migrate into the developing gonad and become gonocytes, which subsequently become germ stem cells, oogonia and spermatogonia depending on the gonadal sex. Oogonia and spermatogonia undergo differentiation through meiosis to produce eggs in the ovary and sperm in the testis [1]. Germ cells maintain totipotency throughout development [2]. Although gametes are highly specialized cells, they demonstrate this potential upon fertilization to generate a new organism [3]. Understanding of germ cell-specific molecular regulators is a key for dissecting the molecular mechanisms underlying germ cell development and totipotency.
Studies in model organisms have identified both evolutionarily conserved and divergent regulators of germ cell development [4], and revealed the importance of transcriptional repression of somatic fates and post-transcriptional control of gene expression in regulating germ cell development [2,5]. Remarkably, many evolutionarily conserved germ genes - including dazl, dead end and vasa - encode RNA-binding proteins. However, how these germ genes are regulated for expression at the transcriptional level has remained essentially unexplored. One exception is the recent identification of microphthalmiaassociated transcription factor (Mitf) as a transcription activator of germ gene expression in medaka [6]. Mitf is a basic helix-loop-helix/ leucine zipper protein and acts as a master regulator of melanocyte development [7,8]. Mitf plays a critical role also in the development of retinal pigment epithelium [9-12] and has been documented in human pigment diseases [13,14].
Retinoic acid (RA) has been demonstrated to play important roles in diverse processes including patterning of the tetrapod limb [15] and the central nervous system [16], formation of blood vessel and bone [17], osteogenesis [18] and specification of cardiomyocytes [19] and liver cells [20]. RA is the active derivative of vitamin A (or retinol), which is synthesized intracellularly and transported to the nucleus where it initiates transcription of target genes [21]. RA functions via binding to retinoic acid receptors (RARs), which bind preferentially as heterodimers with the retinoid X receptor (RXR) to the RA response element (RARE) in the regulatory regions of target genes. The RARE is a direct repeat (DR) of GGTCA spaced by 1~7 nucleotides with TGACC. These receptors are ligand-dependent transcription factors. An RAR:RXR heterodimer binds to the RA-responsive element (RARE) to regulate transcription.
Recent studies have revealed that RA plays essential roles in various processes and aspects of germline development. RA signaling determines the germ cell fate in mice [22]. RA induces the transdifferentiation of myoblasts into premeiotic Stra8-positive cells [23]. More profoundly, RA has been best implicated in controlling meiotic entry. In mice, germ cells within the developing gonad follow a sexually dimorphic pathway. Germ cells in the ovary enter into the meiotic prophase during embryogenesis, whereas germ cells in the testis arrest in G0 of mitotic cell cycle and do not commence meiosis until after birth. Retinoic acid (RA), synthesized from the dietary vitamin retinol or vitamin A, regulates meiotic entry in both male and female germ cells. In female mice, meiosis is initiated soon after gonadal sex determination by 12.5 days post coitum (dpc). This initiation can be blocked by RA antagonists or by disruption of the RA-regulated gene Stra8 [24,25]. In contrast to the fetal ovary, gonocytes in a 12.5-dpc fetal testis enter the G0 phase of the cell cycle and do not divide until after birth. The exit of male gonocytes from the cell cycle is not a consequence of their inability to respond to RA, because the gonocytes of a fetal testis without RA enter meiosis when incubated with RA [22]. It is also not the result of a lack of RA synthesis, as ALDH1A2, the rate-limiting enzyme in RA biosynthesis, is expressed in the mesonephros of both sexes [22]. The testicular RA concentration at 12.5 dpc is only 20% of the ovarian concentration, because the testis expresses CYP26B1, an enzyme that converts RA to inactive metabolites [25]. In fetal Cyp26b1-null mice, the testicular RA level is elevated 3-fold and gonocytes initiate meiosis at 12.5 dpc [26]. Therefore, the RA concentration, but not the genetic sex of the gonocytes, determines the initiation of meiosis in mice.
RA regulates meiotic entry also in chicken [27] and human [28]. The importance of RA signaling in mitotic-to-meiotic transition is well-exemplified by the identification of stra8, one of the genes stimulated by RA, which is a marker of premeiotic mouse germ cells [29,30]. In vivo, RA promotes meiosis via the down-regulation of Nanos2, whereas FGF9 inhibits meiosis through the upregulation of Nanos2 [31]. In vitro, RA promotes proliferation of PGCs from mouse [32] and chicken [33], and induces differentiation of embryonic stem (ES) cells from mouse and fish [34,35].
RA is required also for the maintenance of adult mammalian spermatogenesis. Upon vitamin A withdrawal in adult rodents, germ cells fail to enter into meiosis and remain undifferentiated, and spermatogenesis is disrupted by the loss of differentiated germ cells, which accompanies the accumulation of spermatogonia within the seminiferous epithelium [36]. These results support a notion that RA sets a balance between germ stem cell self-renewal and differentiation via controlling meiotic entry in both oogenesis and spermatogenesis of mammals.
This study was aimed at determining the role of RA in regulating germ gene expression and germ cell development in lower vertebrates. We use the fish medaka (Oryzias latipes) as a model to study stem cells [37-40] germ cells [40-42] and sexual development [43,44]. We have recently identified Mitf as a first transcriptional activator of germ genes [6]. Here we show that RA suppresses Mitf-activated germ gene expression in stem cell cultures and exhausts the spermatogonial stem cell pool in organ culture.
Fish
Work with fish followed the guidelines on the Care and Use of Animals for Scientific Purposes of the National Advisory Committee for Laboratory Animal Research in Singapore and approved by this committee (permit number 27/09). Medaka (Oryzias latipes) strain i1 and transgenic line Vg were maintained under an artificial photoperiod of 14-h/10-h light/darkness at 26°C as described [40,45,46]. In medaka, sex maturation occurs until 3 months post hatching. Breeding males and females measuring ~4 cm in length at 6~7 months were used for organ culture and analysis.
Gene identification and RT-PCR
The medaka RARs and RXRs are annotated in the genome sequence (http://www.ensembl.org/index.html). Total RNA was isolated by using the Trizol reagent (Invitrogen) and used for cDNA synthesis by using oligo(dT)25 and the MMLV reverse transcriptase (Invitrogen) as described [47]. RT-PCR was run in 25-μl volume containing 10 ng of cDNA for 30 cycles (for β-actin) and 35 cycles (RARs and RXRs) of 10 s at 94°C, 15 s at 58°C and 60 s at 72°C. Information on genes and PCR primers used were provided in Table 1. The PCR products were separated on 1.2% agarose gels and documented with a bioimaging system (Synoptics). The PCR products were sequenced by using the BigDye Terminator Cycle Sequencing Ready Reaction Kit and analyzed on the ABI 377 sequencer (Perkin-Elmer, USA).
| Gene | Accession Number | Names and Sequences of Primers | Size (bp) |
|---|---|---|---|
| rarα1 | ENSORLG00000004373 | α1F: AAGTCCTCAGGGTACCACTATGGT α1R:GGCGCTCAGCAGTCCCGTCTCTGC |
720 |
| rarα2 | ENSORLG00000008502 | α2F: ATGTTTGACTGCATGGACGTGCTG α2R:GGATGGGAAAGTTTCTTGATGGGC |
564 |
| rarg1 | ENSORLG00000007861 | g1F: GTACCCAGCTCTCCGTCTCCACCT g1R: CAGCTGAACACGGTGGTCTGAGCT |
450 |
| rarg2 | ENSORLG00000015382 | g2F: CCTCACATGTTTCCCCGCATGCTG g2R:CGCCGCCTTCTTCTGCACGGCCGC |
360 |
| rxrβ | ENSORLG00000007020 | xβF: ATGTCCTCACAGCTCCCTAACAGA xβR:TTTGCCAGAGGAGCGGTCCCCACA |
300 |
| rxrg | ENSORLG00000016690 | xgF: AGTGTCAGCAGCTCCGAGGACATC xgR: ACAGTACTGACAGCGGTTCCGCTG |
270 |
| mitf1 | ENSORLG00000013461 | m1F: GTAGATTACATCCGCAAGCTG m1R: ACAGCCCTGCTCGTTTTCATCCAT |
501 |
| dazl | NM_001104799 | dzF: AATACCTTCATCATCCTCACCTC dzR: AAGTGAAGGAACGTTTGACC |
328 |
| dnd | NM_001164516 | ddF: CAGAGCAAGGTGGTGAACTTG ddR: CACCTTGTAGGTGAAGTACAG |
966 |
| vasa | NM_001104676 | vsF: CGTCCAGCACGTGGTGAACT vsR: TTCCGGACTTGAACGTGGG |
486 |
| β-actin | NM_001104808 | MA1: TTTCAACAGCCCTGCCATGTAC MA2: CCTCCAATCCAGACAGAGTATT |
650 |
1) PCR was run with a common annealing temperature of 58°C for all primers used
Table 1: Primers used for RT-PCR analysis1).
Reporter and effector plasmids
Six luciferase (Luc) reporter plasmids and the effector plasmid pXmitf used in this study have been described [6,48]. pXmitf expresses the Xiphophorus mitfm [49]. Plasmid DNA was prepared by using the Qiagen Midi-preps kit (Qiagen).
Cell Culture and luciferase reporter assay
The medaka ES cell line MES1 and spermatogonial line SG3 were maintained on gelatin-coated tissue culture plastic ware in medium ESM4 [34] and transfected as described [50]. For reporter assays, cells were transfected in 96-well plates as described [6]. In some experiments, cells were treated with all-trans RA (Sigma). For this, RA stock (5 mM) dissolved in dimethyl sulfoxide (DMSO) was added to pre-warmed ESM4 to a final concentration of 2 μM. Controls were treated only with DMSO as vehicle. The RA-containing ESM4 was applied to cells after incubation with the transfection reagent-DNA complex. At 3 or 4 days post transfection, cells were measured for luciferase activity by using the Dual-GloTM Luciferase assay kit (Promega) and measured on GlomaxTM 96 Microplate Luminometer (Promega). The reading from pGL3-basic was considered as background, and that from pTAluc treated as unit activity. Relative luciferase activity from vectors was normalized to that of pTAluc.
Testicular organ culture
Testes were dissected aseptically from 9 Vg adult medaka males. A medaka has two testes, one in the left abdomen and the other in the right abdomen. Intact gonads were seeded in 24-well plates, one testis per well containing 0.5 ml of ESM2 medium with or without 10 μM of RA. Medium ESM2 has proven suitable for deriving and maintaining medaka ES cells [34,35,51,52] and testicular cell lines including SG3 capable of test-tube sperm production [47]. ESM2 is similar in osmolarity to media capable of supporting male germ cell proliferation and differentiation in primary culture of medaka testis [53] and primary spermatocytes [54].
Testicular cryosections
Testes were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) at 4°C for 1 h and subjected to cryosectioning as described [46]. The sections were stained for nuclei with DAPI (1 μg/ml) for 10 min at room temperature, washed three times in PBS and mounted for microscopy [55]. Germ cells were identified by transgenic GFP expression from the germ cell-specific VAS promoter. Different stages of germ cell development in the adult testis were recognized by relative intensities of GFP signal, cellular location and morphology within the ordered testicular structure. A total of 9 pairs of testes were subjected to detailed analyses by sectioning. Three testes were examined at 3, 21 and 28 days of culture in the absence of RA. In parallel, three testes were analyzed at the same intervals of culture in the presence of RA.
Microscopy and photography
Tissue sections and cell cultures were observed under a Zeiss Axiovert2 invert microscope and/or Axiovert200 upright microscope by using a Zeiss AxioCam MRc digital camera [35,55].
Statistics
Statistical analyses were calculated by using Graphad Prism v4.0. Data consolidated were presented as mean ± s.d., and p-values were calculated by using nonparametric Student’s t-test as described [46].
Consensus RARE site present in medaka germ gene promoters
Since RA regulates gene expression through binding to RAREs, we wanted to examine whether the consensus RARE sequence was present in the promoter region of medaka germ genes. We chose dazl, dnd and vasa that have been characterized as germ cell markers [55-59]. In particular, VAS, a 5.1-kb vasa promoter is able to drive germ cell specific GFP expression [58]. There are 1~2 full RARE sites and 7~12 RARE half sites within the 4.2-kb dazl promoter (DAZ), 5.4-kb dnd promoter (DND) and 5.1-kb VAS, respectively (Figure 1A), revealing them as putative RA target genes.
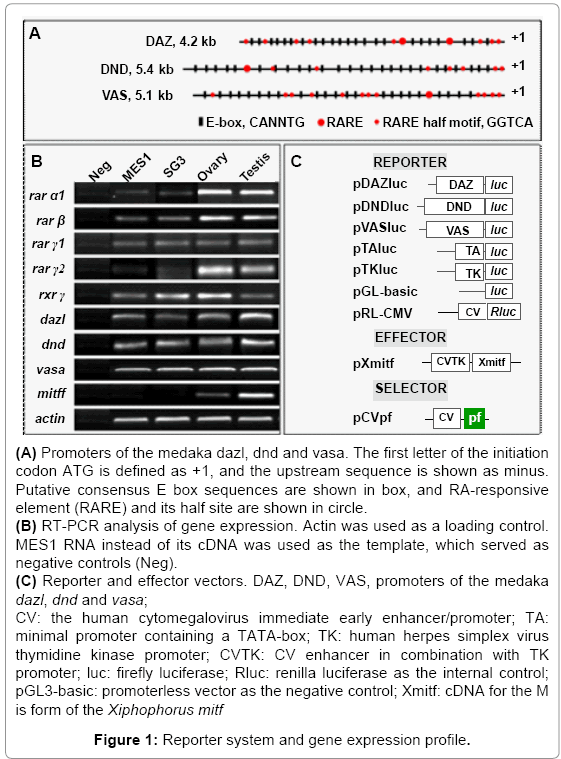
Figure 1: Reporter system and gene expression profile.
Expression of RA signaling genes in medaka stem cells and testes
In mammals, there are three RA receptors (RARα, RARβ and RARγ) and three retinoid X receptors (RXRα, RXRβ and RXRγ) that initiate the signaling through binding by RA. These receptors may exist as duplicate in teleosts due to an ancient whole genome duplication event in the fish lineage. In medaka, four RARs and two RXRs are annotated in the genome (http://www.ensembl.org/index.html). We examined the RNA expression of five of these receptors by RT-PCR analysis. All the 5 genes exhibited detectable expression in the medaka ES cell line MES1, spermatogonial cell line SG3, testis and ovary (Figure 1B). This indicates that the RA signaling pathway is active in medaka ES cells and male germ stem cells in culture as well as the adult gonad.
RA suppresses Mitf-activated germ gene transcription in stem cell cultures
Previously we have shown that Mitf is expressed in medaka germ cells in vivo and capable of activating germ gene transcription in vitro [6], coincident with the presence of multiple putative E-box consensus element, to which Mitf binds to regulate target gene transcription, in the three germ gene promoters (Figure 1A). For reporter assays, these promoters were individually inserted upstream of the fire fly luciferase (luc) in the promoterless construct pGL3-basic, resulting in pDAZluc, pDNDluc and pVASluc respectively (Figure 1C). We made use of SG3 and MES1 cells for transfection and reporter assay. SG3 is a male germ stem cell line from the adult medaka spermatogonia and expresses germ genes dazl and vasa [55]. MES1 is a medaka ES cell line capable of chimera formation [45,46,60]. Both cell lines are suitable to study germ gene expression by reporter assay [6].
In both cell lines, the three germ gene promoters alone exhibited activities approximately 60~120 times the minimal promoter TA (Figures 2A and 2B). We used pXmitf as effector, a construct expressing the Xiphophorus mitfm, because Xmitf has been well-documented [61] and proven to be efficient to induce melanocyte in MES1 [49] and SG3 cells [62]. Upon cotransfection with pXmitf, the activity increased by 3.6, 3.8 and 4.4 fold in SG3 cells (Figure 2A), by 5.9, 6 and 5.5 in MES1 cells (Figure 2B) fold for DAZ, DND and VAS, respectively. When RA was added to cell cultures co-transfected with pXmitf and a reporter plasmid, the transcription activity was reduced by 3~5 fold to the basal level in both SG3 and MES1 (Figures 2A and 2B). Clearly, RA abolishes Mitf-activated germ gene transcription in medaka ES cells and adult male germ cells in culture, suggesting that the three germ genes are targets of the RA signaling.
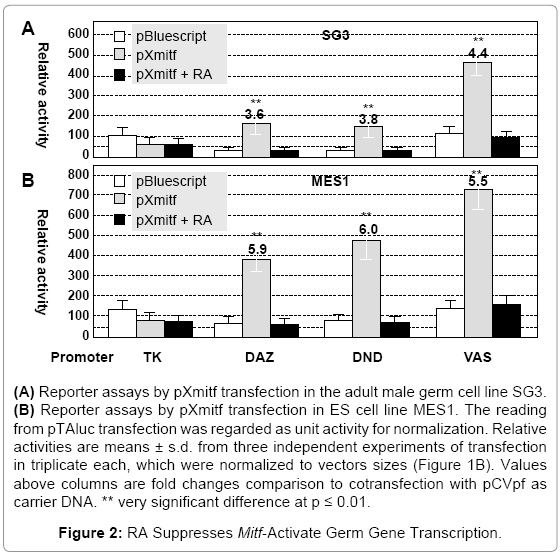
Figure 2: RA Suppresses Mitf-Activate Germ Gene Transcription.
RA abolishes the spermatogonial cell pool
To analyze the effect of RA treatment on adult germ cells, we dissected intact Vg testes and seeded them for organ culture in medium ESM2. The Vg testis shows strong GFP expression in the periphery rich in spermatogonia, spermatogenic cysts but little GFP signal in the efferent duct containing sperm (Figures 3A and 3B). Within the testis under these conditions, spermatogenesis occurs during the first three weeks of organ culture in the absence and presence of RA, because different stages of spermatogenic germ cells, ranging from spermatogonia to motile sperm, were released from the intact testes via the efferent duct (Figures 3C-3F). These results suggest that the adult testis under organ culture conditions used retains the capability of spermatogenesis in vitro.
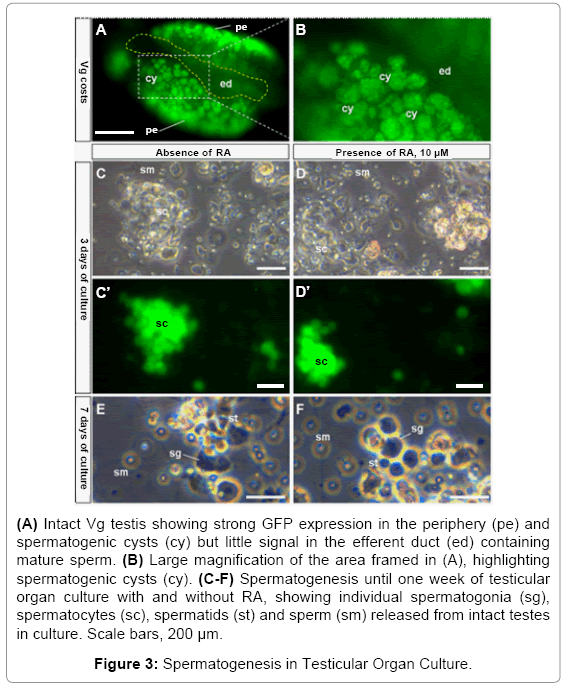
Figure 3: Spermatogenesis in Testicular Organ Culture.
Spermatogenesis diminished gradually and essentially ceased after 3 weeks of organ culture in the presence of RA. Since RA stimulates meiotic entry in chicken and mammals [25,27,28], we examined the testicular structure on cryosections. In medaka, the adult testis consists of cysts of germ cells, each cyst is surrounded by Sertoli cells and comprises of synchronously developing germ cells. Cysts of different spermatogenic stages are arranged in such a way so that the spermatogonial cysts are at the testicular periphery, followed by the cysts of primary spermatocytes, secondary spermatocytes and spermatids, with sperm being matured at the testicular center and stored in the lumen located at the center. Besides a peripheral location, spermatogonia exist as singlets, doublets or small clusters from the periphery towards the center, exhibit relatively weaker nuclear staining and the strongest VAS-driven GFP expression. When spermatogenesis proceeds, the number of male germ cells per cyst increases, nuclear staining becomes more intense and VAS-driven GFP expression decreases (Figures 4A and 4B). These features in combination with the sequential arrangement of different-stages of spermatogenic cysts allow for unambiguous definition of individual spermatogenesis stages, and thus a precise analysis of normal and abnormal spermatogenesis in cultured testes. After 3 days of culture, the control testis maintained in the absence of RA exhibited normal distribution of all stages of spermatogenesis ranging from spermatogonia to sperm (Figures 4A and 4A’. The testis maintained for 3 days in the presence of RA displayed a similar structure but a remarkable reduction in the number of spermatogonia (compare figures 4B and 4B’ with figures 4A and 4A’. Thus, in short-term testicular culture, RA reduces the number of spermatogonia.
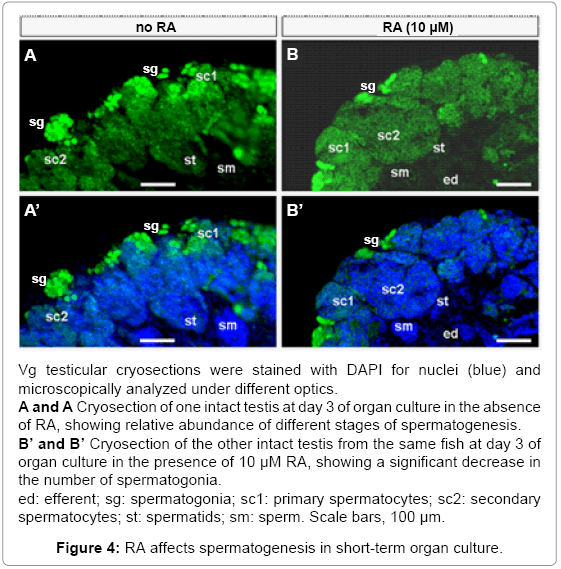
Figure 4: RA affects spermatogenesis in short-term organ culture.
After 21 days of culture in the absence of RA, the testis still exhibited many spermatogonia and early meiotic germ cells (e.g. primary spermatocytes), which distributed mostly in the peripheral region, as well as late stages of spermatogenic cells (spermatids and sperm), which were found outside the peripheral region (Figures 5A- 5D). In contrast, after 21 days of culture in the presence of RA, the testis became completely exhausted of spermatogonia, leading to a peripheral region essentially free of GFP-positive germ cells (Figures 5E and 5F). After 28 days of RA treatment, the number of germ cells was remarkably reduced, leading to areas of merely somatic cells not only in the peripheral but also central regions of the testis (Supplementary Figure S1). Taken together, RA treatment in extended organ culture disrupts the maintenance of spermatogonial stem cell pool.
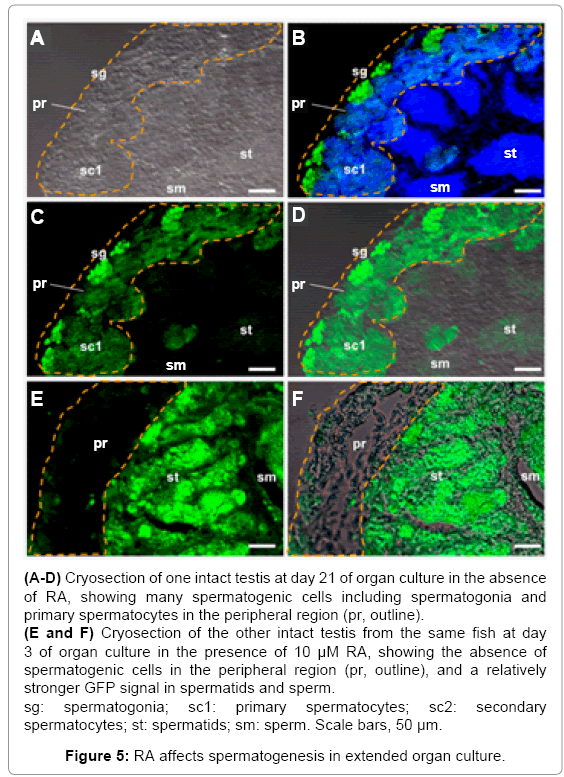
Figure 5: RA affects spermatogenesis in extended organ culture.
RA exerts different effects on germ gene expression depending on stages
The fact that RA inhibits Mitf-mediated germ gene expression in cell culture provoked us to examine its effect on testicular germ gene expression in organ culture. Compared to the control testis maintained in the absence of RA (Figure 4A), the intensity of GFP signal altered markedly after 3 days of RA treatment: It became weaker in spermatogonia but stronger in meiotic cells (primary and secondary spermatocytes; Figure 4B). Elevated expression in late meiotic cells (spermatids) was most evident in the testes following RA treatment for 21 days (Compare Figures 5C and 5E). Furthermore, RA treatment prevented the loss of GFP expression in sperm (Compare Figures 5C and 5E). Taken together, VAS-driven transgenic gene expression is suppressed in spermatogonial stem cells but elevated in meiotic and post-meiotic male germ cells by RA treatment in testicular organ culture, suggesting that RA exerts different effects on germ gene expression depending on stages of germ cell development.
In mammals, the regulatory role of RA has recently been well documented in germline development, ranging from fate decision over proliferation to meiosis entry [22,25,27,28,32,33]. In this study, we provide first evidence that RA regulates germ gene expression and germ cell development also in medaka as a lower vertebrate model, suggesting that the RA signaling plays a highly conserved role in controlling germline development from fish to mammals.
In vitro in the absence of somatic cells, RA promotes proliferation of PGC cultures from mouse [32] and chicken [33]; in vivo, RA stimulates differentiation and inhibits self-renewal of gonadal germ stem cells of mouse [31], human [28] and chicken [27]. The mechanism underlying the effect of RA on proliferation in vitro or differentiation in vivo of germ stem cells remains unknown. A difference in response to RA treatment might be due to a difference between PGCs and spermatogonia in nature and assay condition. In this study, the testicular organ culture system reveals that RA stimulates differentiation and inhibits selfrenewal of spermatogonial stem cells. This suggests that male germ cells within dissected testes in organ culture resemble those of intact testes in live animals in response to RA treatment, underscoring the usefulness of organ culture system for studying germ cell proliferation and differentiation.
In this study, we show that all the three germ genes possess the RA responsive element in their regulatory regions and that, more importantly, RA abolishes activation of the germ genes’ transcription by Mitf. These results suggest that RA signaling may be directly involved in regulating germ gene transcription. Since our reporter assay has used stem cell culture and RA is a strong inducer of stem cell differentiation in medaka [34,35], it is also likely that RA may prevent Mitf-activated germ gene expression through induced differentiation of stem cells. RA works through binding to its receptors. We reveal that RA receptors are indeed expressed in both cell lines used for reporter assays. Future work will determine whether RA receptors are differentially expressed at different stages of medaka germ cell development of both sexes and required for the regulation of germ gene expression.
These results show that transcription of germ genes in cell culture is controlled by two opposing mechanisms, one is activation by Mitf and the other is silencing by RA. Furthermore, the negative transcriptional effect of RA is dominant over the activating effect of Mitf. A similar observation has also been made with the mouse oct4, an octamerbinding transcription factor that plays a key role in the maintenance of pluripotency and exhibits pluripotency-specific expression in ES cells and germline [63]. Oct4 is down-regulated by RA via several RARE half-sites in the proximal promoter [64].
We observe that RA does reduce VAS-driven GFP reporter expression in spermatogonia in testicular organ culture, which is in accordance with RA-mediated suppression of Mitf-activated germ gene expression in stem cell culture. It has been shown that endogenous expression occurs at the highest level in spermatogonia, declines remarkably during meiosis and becomes hardly detectable in postmeiotic stages for germ genes examined in this study, namely dazl and vasa [55], dnd [56] and mit [6].
In medaka, it has been reported that expression of the vasa RNA is at the highest level in spermatogonia and reduced in meiosis and post-meiotic phase on testicular sections [55], and that vasa promoterdriven GFP expression occurs at the highest level also in mitotic and early meiotic germ cells in the peripheral cysts of the whole testis [65]. Results in this study collaborate and extend these reports by revealing that on testicular sections, VAS-driven GFP expression is the highest in spermatogonia, remains abundant in primary spermatocyte and reduces in meiotic and post-meiotic germ cells.
An interesting observation obtained in this study is that RA reduces GFP reporter expression from the vasa promoter in spermatogonial cells but enhances GFP expression in meiotic and post-meiotic male germ cells. This finding is consistent with the reports that vasa is essential for proliferation, differentiation and meiotic progress of germ cells in mouse [66] and human [67]. In mice, it has been reported that RA as well as stem cell factor increases meiotic entry, upregulates expression of meiotic genes stra8 (master regulator of meiosis), dmc1 (early meiotic marker essential for meiotic recombination), bmp6, btg4, dzip1l and phf7, and downregulates expression of stemness genes (pcgf2, pcgf6 and numb) and spermatogonial markers (egr2, pole, ptn, tex16 and zfpm2), which are known to turn off at the onset of meiosis [68]. Most recently, the medaka 5.1-kb VAS promoter used for driving GFP expression has been reported to contain up to 11 positive and negative regulatory regions, which exert opposing mechanisms underlying vasa transcription [48]. This study identifies one RARE and 12 copies of the RARE half site. Future work is needed to determine the precise region(s) on which RA acts to regulate stage-specific vasa expression.
We thank Jiaorong Deng for fish breeding, Dr. J. Altschmied (Düsseldorf, Germany) for kindly providing plasmid pXmitf, and Yovita Ida Purwanti for part of work. This research was supported by the National Research Foundation of Singapore (NRF-CRP7-2010-03 and NRF-CRP 002-082).
