Review Article - (2012) Volume 1, Issue 1
Molecules are rapidly replacing morphology as the preferred source of evidence for generating phylogenetic hypotheses. Critics of morphology claim that most morphology-based characters are ambiguous, subjective and prone to homoplasy. In this paper we summarize the results of recent Bayesian and parsimony-based cladistic analyses of the gross muscle morphology of primates and of other animals that show that morphological evidence such as muscle-based data is as capable of recovering phylogenies as are molecular data. We also suggest that recent investigations of neural crest cells and muscle connectivity might help to explain why muscles provide particularly useful characters for inferring phylogenies. Lastly, we show how the inclusion of soft tissue-based information in phylogenetic investigations allows researchers to address evolutionary questions that are not tractable using molecular evidence alone, including questions about the evolution of our closest living relatives and of our own clade.
Keywords: Muscles; Molecules; Morphology; Primates; Human evolution; Phylogeny; Evolutionary biology
There has been a marked decline in the number of morphologybased phylogeny reconstructions. Critics of the use of morphology claim that morphological characters are ambiguous, that the delineation of homology is subjective, and that phenotypic features are particularly prone to homoplasy [1-5]. In this paper, we use our experience with the morphology of a wide range of animal groups, but most recently with the morphology of the striated muscles of primates, as well as with other areas of knowledge such as developmental and evolutionary biology, to argue that that morphological evidence such as muscle-based data is as capable of recovering phylogenies as are molecular data. We briefly review the results of recent Bayesian and parsimony analyses of myological characters that addressed the higherlevel phylogeny of the whole primate clade, plus the results of analyses that used muscle morphology to recover the relationships among other groups of animals. We also review research on neural crest cells and muscle connectivity that might help to explain why muscles are apparently particularly useful for inferring phylogeny. We then address the phylogenetic and evolutionary implications of the data discussed in the paper in terms of the role myology can play in evolutionary biology and systematics.
Molecular evidence from whole protein studies [6], singlecopy DNA-DNA hybridization [7,8], nuclear DNA sequences [9- 13], mitochondrial DNA sequences [14-16], from both nuclear and mitochondrial DNA sequences [17,18] and also from Alu elements [19] provide support for the relationships among the major primate clades shown in Figure 1. It should be noted that although a few molecular studies have contradicted some of the higher (more inclusive) clades shown in the tree of Figure 1 (e.g., Chatterjee et al. supported a Tarsius + Strepsirrhini group) [20], the clades depicted have, in general, being consistently supported in the most comprehensive molecular phylogenetic analyses is a MRP - matrix representation with parsimony - supertree of mammals [17,18,21,22]. With respect to the more specific intra-relationships (within each of these higher clades) that are shown in Figure 1, the only major difference among the results of the more recent analyses is that in Fabre et al.’s and in Perelman et al.’s[17,21] studies some cebids (e.g., Saimiri or Callithrix) [17,21] appear closer to aotids (e.g. Aotus) than to other cebids (e.g., to Callithrix in Fabre et al., or to Saimiri in Perelman et al.) [17,21], while in Arnold et al. [18] study the cebids form a monophyletic group. The close relationship between Callithrix and Saimiri, and the monophyly of the Cebidae proposed by Arnold et al. [18] are consistent with the results of the most recent and complete molecular analysis of platyrrhine relationships [23-27]. Thus, it can thus be said that the tree shown in Figure 1 summarizes the relationships among the major groups within the order Primates that are supported by the most up-to-date molecular evidence, and it is this tree that we can thus be used to validate other non-molecular methods for recovering phylogeny.
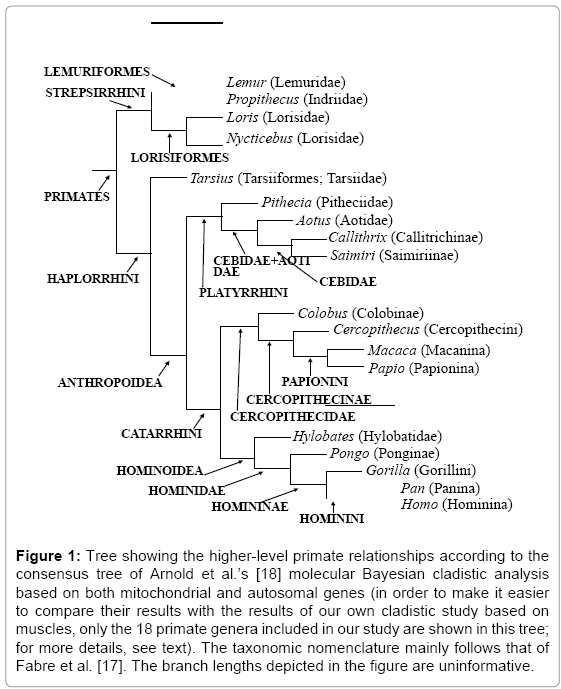
Figure 1: Tree showing the higher-level primate relationships according to the consensus tree of Arnold et al.’s [18] molecular Bayesian cladistic analysis based on both mitochondrial and autosomal genes (in order to make it easier to compare their results with the results of our own cladistic study based on muscles, only the 18 primate genera included in our study are shown in this tree; for more details, see text). The taxonomic nomenclature mainly follows that of Fabre et al. [17]. The branch lengths depicted in the figure are uninformative.
The results of the first comprehensive cladistic analysis of the higher-level phylogeny of all of the major extent groups of primates (18 genera) that included both molecular characters and a large number of morphology-based characters [28] was consistent with the tree shown in Figure 1. The 264 morphology-based characters (mainly compiled from previous data) [29,30] used by Shoshani et al. [28] included some soft-tissue data, but the vast majority of the characters were based on the hard tissue anatomy of various regions of the body. Although Shoshani et al. [28] stressed that their study was the first published report “based on a rigorous maximum parsimony computer analysis of a large data matrix on living Primates” to provide “morphological (cladistic) evidence” for the chimp-human clade, that clade structure was only weakly supported (e.g., their cladistic analysis had a bootstrap support value of just 42 (out of 100).” In particular, the results of the Shoshani et al. [28] analysis supported the monophyly of the clade Haplorrhini (i.e., Tarsius plus anthropoids), as most (but not all) molecular studies do (Figure 1) but Shoshani et al.[28] noted that various paleontological studies have supported a Tarsius-Strepsirrhini clade. This is just one of several examples that are cited to demonstrate that the results of fossil-based analyses are contradicted by molecular and neontological morphological evidence, but according to Shoshani et al. [28] in this case the discrepancy may be due to an incorrect interpretation of the paleontological evidence (e.g., some of the features considered by paleontologists to support a closer relationship between tarsiers and strepsirhines are probably plesiomorphies). But does this mean that molecular data should have primacy over morphological data with respect to phylogeny reconstruction? To address this question, in this paper we thus focus on how evidence about muscle morphology has contributed to phylogeny reconstruction in various groups of animals.
In one of the first studies that considered the utility of myological data for phylogenetic reconstruction, Borden [31] described the configuration and variation of 93 muscles in 15 species of the genus Naso or Unicornfishes (Teleostei: Percomorpha) and discussed the phylogenetic implications of the results. Borden suggests [31] that phylogenetic studies have neglected evidence from myology because “investigators may be reluctant to use myology due, for example, to the plethora of names that have been used to describe the same muscles, to the realization that osteological proficiency is mandatory in order to identify muscles, leading them to concentrate only on osteology, or to the requirement of potentially finer dissection to preserve muscle bundles and nerves”. In consequence “of those studies using myology as a basis of information, most are functional works often analyzing the role of various muscles in feeding or locomotion or comparing a muscle or specific group across a number of taxa systematically and/ or ecologically related” [31]. Diogo [32,33] compared the incidence of homoplasy and the utility of 91 myological and 303 osteological characters used in the reconstruction of the higher-level phylogeny of a diverse group of teleosts, the Siluriformes (or catfish). The results of both of these studies indicate that osteological structures generally display more morphological variation (i.e., incorporate more character states) than do myological ones. Thus, although hard-tissues usually provide more characters suitable for phylogenetic analyses, myology-based characters are generally more effective at recovering the relationships among higher clades that are supported by molecular data. Diogo [34] increased the scope (a total of 356 characters in 80 extant and fossil terminal taxa) and reach of his cladistic analyses by extending them to include the osteichthyan clade (bony ‘fish’ plus tetrapods), and he also found that hard tissue structures (bones and cartilages) displayed more variation than the myological ones. For example, the 81 osteological structures examined for Diogo’s [34] cladistic analysis provided 198 phylogenetic characters (i.e., 2.4 phylogenetic characters per osteological component), while the 63 muscles examined provided 122 phylogenetic characters (i.e., 1.9 phylogenetic characters per muscle). However, the mean Retention Index (RI) of the informative muscular characters examined by Diogo [34] (0.82) was higher than that of the informative osteological characters examined (0.71) (i.e., the myological characters used by Diogo [34] were on average more useful for the retention of the clades obtained in the cladistic analysis of his complete dataset than the hard tissue-based characters). A similar pattern was seen for the Consistency Index (CI), in which the number of informative myological characters used by Diogo was significantly higher than that for the informative osteological characters (0.71 and 0.52, respectively). Thus, both of these studies suggest that although osteological structures provide more potential characters for phylogenetic analyses, for one reason or other myological characters are more useful for inferring the phylogenetic relationships among higher clades. This suggestion has been corroborated in studies that have focused on other major vertebrate groups such as teleosts [35,36], birds (e.g., McKitrick [37]), squamates (e.g., Abdala and Moro [38,39]), hominoids (Gibbs (1999); Gibbs et al. (2000); Gibbs et al. (2002) [40- 42]), including fossil groups such as dinosaurs (e.g., Dilkes [43]).
Gibbs et al. [42] suggested that the apparently high reliability of muscle characters for recovering the phylogeny of higher taxa may be due to the way muscles develop. The results of experiments using rhombomeric quail-to-chick grafts to investigate the influence of hindbrain segmentation on craniofacial patterning [44] indicated that rhombomeric populations remain coherent during ontogeny, with rhombomere-specific matching of muscle connective tissue with their attachment sites for all branchial and tongue muscles. However, the Köntges and Lumsden [44] study concerns only the head muscles and it is related to the connective tissue/fasciae associated with the muscles, and not with the ontogenetic and/or phylogenetic origin of these muscles. So, for example, the avian hyobranchialis (‘branchiomandibularis’) is a branchial muscle [45,46] but anteriorly it is attached to hyoid (2nd arch) crest-derived skeletal domains (i.e., the retroarticular process of the mandible) because the anterior part of this muscle is associated with connective tissue/fasciae that is derived from hyoid crest cells. The hyobranchialis is the only muscle studied by Köntges and Lumsden [44] that derives its connective tissue from more than one branchial arch for its posterior moiety (i.e., the 3rd and 4th arches) and, accordingly, it inserts onto 3rd and 4th arch crest-derived skeletal domains. Three hypobranchial muscles, hyoglossus, hypoglossus and genioglossus, are also consistent with the model proposed by Köntges and Lumsden [44]. Previous mapping studies have shown that the myocytes and the innervation of these three muscles are derived from the posterior axial levels of the first somites. However, as explained by Köntges and Lumsden [44] the skeletal attachment fasciae of these three muscles are “derived from the more anterior axial levels of cranial neural crests” and these author’s suggest that this is why the genioglossus and hypoglossus are attached to skeletal elements such as the paraglossals and the ventral basihyoid (Köntges and Lumsden [44]), which are derived from mandibular arch crest originating in the posterior midbrain. It is also why the hyoglossus (‘ceratoglossus’), which is also ontogenetically and phylogenetically derived from the geniohyoideus, is attached to hyoid (2nd arch) crest-derived skeletal elements. The attachments of these three hypobranchial muscles are primarily determined by the origin of the connective tissues/fasciae to which they are associated, and not by their ontogenetic and phylogenetic origin. There are, however, exceptions to the model proposed by Köntges and Lumsden [44]. For example, some mammalian facial muscles that are derived from the second (hyoid) arch and which are apparently associated with connective tissue/fascia also derived from this arch, move into midfacial and jaw territories populated only by frontonasal and first arch crest cells [47-53] have shown that these facial muscles behave, in terms of C-met mutations, as hypaxial migratory muscles. Contrary to most other head muscles, with the exception of the hypobranchial muscles [46] the facial muscles are absent in organisms with C-met mutations, thus suggesting that during ‘normal’ ontogeny these mammalian muscles migrate far away from their primary origin. As noted by Gibbs et al. [42], if the Köntges and Lumsden [44] model “operates elsewhere in the body, it would help explain how muscle gross morphology is conserved, whereas the shapes of the skeletal elements to which the muscles are attached are susceptible to changes that contrive to obscure phylogeny”. With regard, at least, to teleost fishes, the principal points of muscular origin and insertion do seem to be relatively stable [32].
Another contributory factor suggested by Gibbs et al. [42] phenotypes” because “whereas the mass of a muscle may be affected by activity or inactivity, its attachments are unlikely to be” [42]. However, homoiology cannot be the whole explanation for the difference in phylogenetic reliability between osteological and myological structures, since dental enamel, for example, does not remodel and it therefore not obviously subject to homoiology [42]. But other authors have suggested that functional or developmental constraints may result in tooth morphology being particularly prone to homoplasy, and, therefore, dental structures may be a poor source of evidence for phylogenetic reconstructions [54].
Bayesian and Parsimony Analyses of Primates Based on Muscles
Soft tissue data have previously been incorporated into some morphology-based investigations of the relationships among the taxa within the primate clade [28,29] but except for Gibbs et al.’s [41,42] study, soft-tissue characters have always been substantially outnumbered by those based on hard tissues. This near total reliance on osteological data is particularly unfortunate because it leads researchers to equate ‘morphology’ with ‘hard-tissue’ morphology. For instance, Grehan and Schwartz [55] have recently argued that the results of their cladistic analysis shows that, contrary to molecular evidence, ‘morphology’ strongly supports a (Pongo, Homo) clade. However, their analysis of ‘morphology’ only included three myological characters.
Gibbs et al. [42] reported the results of a phylogenetic metanalysis of information about the soft tissue morphology of the great and lesser apes. Of the soft tissue structures in the 6th edition of the Nomina Anatomica [56], information from the literature was available for at least one of the apes for 621 out of the 1783 (i.e., c.35%) listed, but only 240 of the listed structures were found to have published information for all four of the non-human anthropoid apes. To be useful for a phylogenetic analysis, more than one state of a structure must exist and one of those character states must be present in two or more of the apes; these additional criteria reduced the character count to 171. These 171 structures were themselves a biased sample of the soft tissues for muscles (64% of the total) and the limbs (82% of the total) were over-represented. But either because of, or in spite of, these biases the 171 soft tissue structures were effective at recovering a hypothesis of relationships (((Pan, Homo) Gorilla) Pongo) among the hominids that was, and is still, consistent with the consensus hypothesis of relationships supported by most molecular studies and a few cladistic studies based solely or mainly [28] on osteological data (Figure 1). As stressed by Gibbs et al. [42] various hard tissue-based studies have supported a range of different hypotheses, including ones in which modern humans are more closely related to gorillas [57-60] or to orangutans [55,61-63] than to chimpanzees.
A recent analysis [64] attempted to overcome some of the limitations (e.g., lack of visual verification of the data, inter-observer error and narrow taxonomic scope) of the Gibbs et al. [41,42] study. It consisted of a systematic study of the gross anatomy of the muscles of the head, neck, pectoral region and the upper limb across the whole of the primate clade, plus several outgroups. We believe that strength of the Diogo and Wood [64] cladistic analysis is that it explicitly avoided using an arbitrary selection of characters or characters. The only bias in our character selection was the intentional one that we used as our evidence the gross morphology of all of the striated muscles in the regions set out above; we were careful not to cherry-pick that evidence for characters whose distribution was consistent with a preferred hypothesis. We combined data from our own dissections with carefully validated information from the literature and we used parsimony and Bayesian methods to test if the relationships supported by muscles were consistent with the evolutionary molecular tree shown in Figure 1.
The most parsimonious tree obtained from our analysis of 166 head, neck, pectoral and upper limb muscle characters in 18 primate genera and in representatives of the Scandentia, Dermoptera and Rodentia (Figure 2) was 100% congruent with the evolutionary molecular tree (Figure 1). The full list of characters used in the cladistic analyses and of the synapomorphies/apomorphies of each clade/ terminal taxon shown in Figure 2 is given in Diogo and Wood [64,65]. These characters mainly concern the presence/absence of muscles and of muscle bundles and differences in the origin or insertion and in the innervation of these muscle structures; information about the outgroup genera Tupaia (Scandentia), Cynocephalus (Dermoptera) and Rattus (Rodentia), as well as of various other mammalian taxa described in the literature and/or previously dissected by us, was used to polarize the primate character states [65]. Most primate clades shown in Figure 2 are supported by high parsimony bootstrap support values (BSVs) and/or high Bayesian credibility support values (CSVs) (e.g., 8 (47%) of them have BSVs and/or CSVs that are ≥ 94). This is thus the first morphological cladistic study based on a large data matrix to provide compelling levels of support for the chimp-human clade (BSV 75, CSV 94) for, as explained above, in Shoshani et al.’s [28] cladistic analysis including 18 extant primate genera and 264 (mostly osteological) characters, the chimp-human clade had a low support (BSV of 42). When we ran separate cladistic analyses of datasets based on the two main anatomical regions we sampled, namely the head and neck (HN; chars. 1-67) and the pectoral region and the upper limb (PU; chars. 68-166), we found that HN muscles are more effective at recovering the molecular evolutionary tree of primates shown in Figure 1. For example, whereas the consensus tree obtained from the parsimony analysis of 67 HN characters recovered 17 of the 20 clades shown in the parsimony tree of Figure 2, the consensus tree obtained from the parsimony analysis of a larger number (i.e., 99) of PU characters only recovered 12 of the 20 clades. However, despite recovering a smaller number of the clades of the molecular tree of Figure 1 than do the HN characters, the PU muscle characters are particularly effective at recovering relationships at the base of the primate clade (e.g., order Primates and suborder Strepsirrhini). Within both the HN and PU datasets the number of total evolutionary changes per muscle and the frequency of non-homoplastic transitions are similar. This result is consistent with a t-test of variable character transition rates obtained from the Bayesian gamma model. A recent analysis of osteological data revealed that the levels of homoplasy found in the dentition, the cranium, and the postcranium of primates are similar. It is noteworthy, however, that although in our parsimony analysis the frequency of non-homoplastic changes is much the same within the HN and PU datasets (about two-thirds of the changes are non-homoplastic in both datasets), the frequency of reversions within the HN dataset (i.e., 18%, with a ratio of 0.30 reversions per muscle studied) is twice that within the PU dataset (i.e., 9%, with a ratio of 0.16 reversions per muscle).
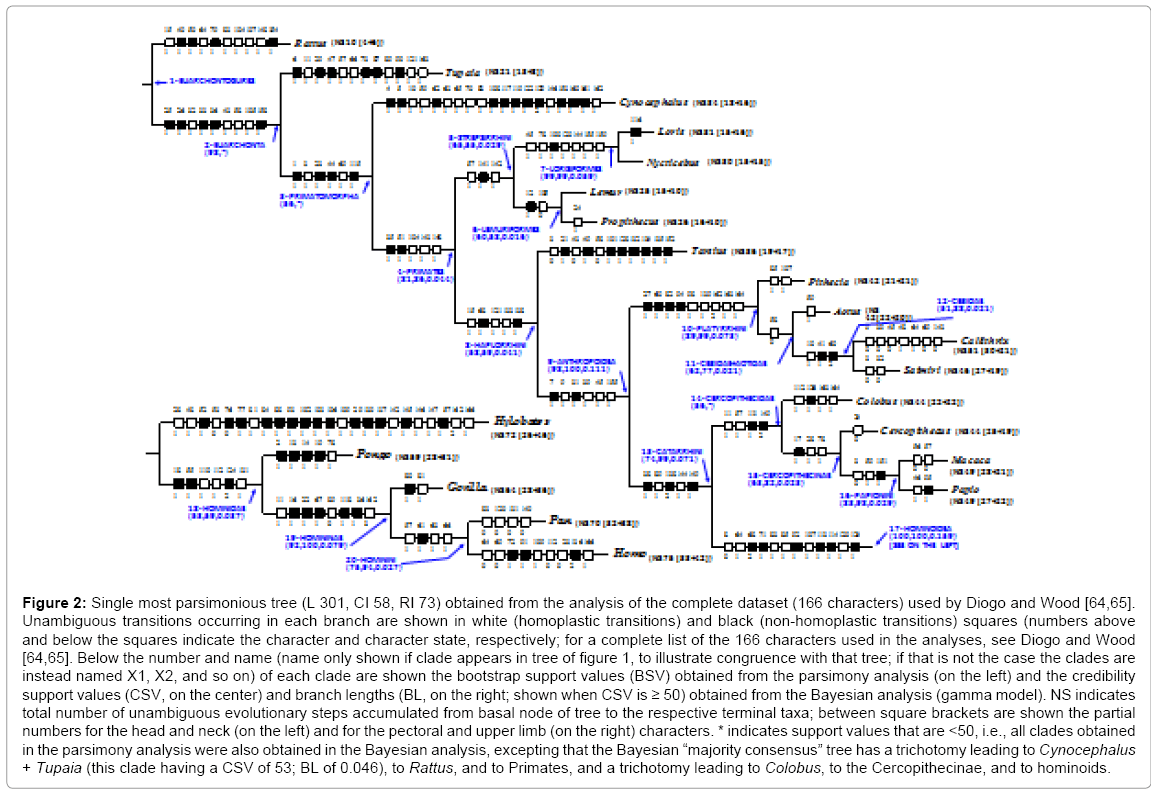
Figure 2: Single most parsimonious tree (L 301, CI 58, RI 73) obtained from the analysis of the complete dataset (166 characters) used by Diogo and Wood [64,65]. Unambiguous transitions occurring in each branch are shown in white (homoplastic transitions) and black (non-homoplastic transitions) squares (numbers above and below the squares indicate the character and character state, respectively; for a complete list of the 166 characters used in the analyses, see Diogo and Wood [64,65]. Below the number and name (name only shown if clade appears in tree of figure 1, to illustrate congruence with that tree; if that is not the case the clades are instead named X1, X2, and so on) of each clade are shown the bootstrap support values (BSV) obtained from the parsimony analysis (on the left) and the credibility support values (CSV, on the center) and branch lengths (BL, on the right; shown when CSV is ≥ 50) obtained from the Bayesian analysis (gamma model). NS indicates total number of unambiguous evolutionary steps accumulated from basal node of tree to the respective terminal taxa; between square brackets are shown the partial numbers for the head and neck (on the left) and for the pectoral and upper limb (on the right) characters. * indicates support values that are <50, i.e., all clades obtained in the parsimony analysis were also obtained in the Bayesian analysis, excepting that the Bayesian “majority consensus” tree has a trichotomy leading to Cynocephalus + Tupaia (this clade having a CSV of 53; BL of 0.046), to Rattus, and to Primates, and a trichotomy leading to Colobus, to the Cercopithecinae, and to hominoids.
Morphological cladistic analyses such as these allow us to address other macroevolutionary topics. For instance, we recently discussed the impact of muscle variations in evolutionary and phylogenetic analyses and in particular the validity of Dollo’s law and the notion of atavism in evolutionary and developmental biology [66]. Another example concerns Bakewell et al. [67] statement that their molecular studies show that “in sharp contrast to common belief, there were more adaptive genetic changes during chimp evolution than during human evolution” and they claim their analysis “suggests more unidentified phenotypic adaptations in chimps than in humans”. The results of the parsimony and Bayesian analyses we refer to above indicate that, at least regarding to the gross morphology of the HN and PU muscles, since the Pan/Homo split the Hominina clade has evolved faster than the panin clade (2.3 times faster according to the lengths of the branches leading to modern humans (9) and to chimpanzees (4) in the parsimony tree of Figure 2 and 2.4 times faster according to the number of changes in the branches leading to modern humans (0.071 changes per character) and to chimpanzees (0.030 changes per character) in the consensus tree obtained from the Bayesian analysis of the complete dataset). In turn, since the split between Gorilla and the Hominini, gorillas have only accumulated two unambiguous muscular apomorphies, whereas there are respectively 8 (4 + 4) and 13 (4 + 9) unambiguous apomorphies leading to extant chimpanzees and to modern humans (Figure 2) (since this split, the branch lengths leading to Gorilla, Pan and Homo in the consensus tree obtained from the Bayesian analysis of the complete dataset are 0.018, 0.057 and 0.098, respectively).
In terms of their significance for our understanding of human evolution, the results obtained from our recent cladistic analyses and comparative anatomical studies seem paradoxical. On the one hand the cladistic analyses suggest there are more unambiguous evolutionary steps (NS) from the base of the tree to modern humans than to any other taxon included in the study (Figure 2). But, on the other hand, our comparative anatomical studies show that modern humans have fewer muscles than most other primates, in particular fewer than in strepsirrhines and tarsiiforms (Table 1). For instance, Nycticebus has a NS of 30 and a range of 133-139 head, neck, pectoral and upper limb muscles in total, while chimpanzees have an NS of 70 and 126 muscles in equivalent regions and modern humans have an NS of 75 but only 123 muscles in total.
| Lemur | Propithecus | Loris | Nycticebus | Tarsius | Pithecia | Aotus | Saimiri | Callithrix | Colobus | Cercopithecus | Papio | Macaca | Hylobates | Pongo | Gorilla | Pan | Homo | |
| Mandibular muscles | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 7-8 | 7-8 | 8 | 8 | 8 | 7 | 8 | 8 | 8 |
| Hyoid muscles (not extrinsic ear) | 25 | 24 | 24-26 | 26 | 24 | 22 | 23 | 21 | 22 | 24-25 | 26-27 | 25-26 | 26 | 26 | 26 | 26 | 26 | 27 |
| Branchial muscles | 14-16 | 14-16 | 15-17 | 14-17 | 16-17 | 14-16 | 14-16 | 15-16 | 14-16 | 13-14 | 16 | 14-15 | 16 | 17 | 14-15 | 15-16 | 15 | 16 |
| Hypobranchial muscles | 12 | 12 | 12-15 | 12-15 | 12 | 12-13 | 11-12 | 12 | 13 | 12 | 12 | 13 | 13 | 13 | 12-13 | 13 | 13 | 13 |
| Pectoral muscles | 17 | 15-16 | 16 | 16 | 17 | 15 | 16 | 16 | 17 | 16 | 17 | 17 | 17 | 14 | 15 | 14 | 14 | 14 |
| Arm muscles | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 4 |
| Forearm muscles | 19 | 19 | 18 | 18 | 19 | 19 | 19 | 19 | 19 | 19 | 19 | 19 | 19 | 19 | 18 | 18 | 19 | 20 |
| Hand muscles | 30 | 30 | 30 | 34 | 32-36 | 22 | 22 | 22 | 21 | 27 | 27 | 27 | 27 | 27 | 20 | 20 | 26 | 21 |
| Total number of muscles | 130-132 | 127-130 | 128-135 | 133-139 | 133-138 | 117-120 | 118-121 | 118-119 | 119-121 | 123-126 | 129-131 | 128-130 | 131 | 129 | 117-119 | 119-120 | 126 | 123 |
Table 1: Table summarizing the total number of mandibular, hyoid (not including the small facial, extrinsic muscles of the ear), branchial, hypobranchial, pectoral, arm, forearm and hand muscles in adults of the primate genera included in our cladistic analyses. Data are from evidence provided by our own dissections and comparisons and from a review of the literature (Diogo and Wood [64,65]); note that in some cases there are insufficient data to clarify whether a particular muscle is usually present, or not, in a taxon (e.g., the number of branchial muscles of Gorilla is given as 15 to 16 because it is not clear if the salpingopharyngeus is usually present, or not, as a distinct muscle in the members of this genus).
As Gould [68] noted in ‘The Structure of Evolutionary discourses on the importance and frequency of ‘progressive evolutionary trends’ have consumed a substantial part of research on the history of clades. However, the importance given to these ‘trends’ bears no necessary relationship with the relative frequency or causal weight of this phenomenon in the natural history of these clades. It seems more related to the general tendency to use ‘progressive trends’ to tell stories. Gould [68] stresses that evolution is a narrative science and he noted that “Western tradition has always favored directional tales of conquest and valor while experiencing great discomfort with the aimless undirected evolution.” He suggested that the focus on ‘progressive evolutionary trends’ is accompanied by the historical under-reporting of examples of undirected evolution. As noted by Gould [68], this type of historical bias is often seen in palaeontological publications, in which examples of stasis are often either non-reported or under-reported because such stability represents ‘no data’. Gould drew an analogy between these biases and “Cordelia’s dilemma”. Cordelia is “King Lear’s honest but rejected daughter”, who, “when asked by Lear for a fulsome protestation of love in order to secure her inheritance” chose to say nothing for she knew that “my love’s more ponderous than my tongue.” Lear, however, mistook her silence for hatred or indifference, and cut her off entirely (with tragic consequences that were later manifest in his own madness, blindness, and death) proclaiming that “nothing will come of nothing.” The equivalent of Cordelia’s dilemma in science is when a signal from nature is either not seen, or not reported. “Most clades, while waxing and waning in species diversity through time, show no overall directionality, but the bias against reporting the existence of such clades means that researchers chronically underestimate the frequency of clades that change all the time but ‘go’ nowhere” in particular during their evolutionary peregrinations.” The results of our recent study support Gould’s contention in the sense that there is no general trend to increase the number of muscles leading to hominoids and to modern humans (Table 1). That is, with respect to the muscles in the regions we have investigated, although modern humans accumulated more evolutionary transitions than the other primates included in the cladistic study (Figure 2) these were evolutionary transitions that did not result in more muscles or muscle components [46,64,65,69-73]. For example, although some of the nine modern human apomorphies acquired since the Pan/Homo split (Figure 2) involve the differentiation of new muscles (rhomboideus major and rhomboideus minor, extensor pollicis brevis and flexor pollicis longus), others involve the loss of muscles (levator claviculae [72] and dorsoepitrochlearis;) [64,65].
We suggest that morphology-based phylogeny reconstructions such as those based on myology should be actively promoted so that they can complement the information obtained in molecular phylogenies. Researchers should use as many different types of data as possible (e.g., muscles, ligaments, bones, cartilages, nuclear DNA, mitochondrial DNA, proteins, Alu insertions, and behavioral and ecological information) for these efforts.
Muscles and other soft tissue data have been particularly neglected in systematics, but the few cladistic analyses based on soft tissues that have been published to date have shown that these tissues can be particularly useful for inferring phylogenetic relationships, including those among fossil taxa. The inclusion of soft tissue-based information in phylogenetic investigations allows researchers to address evolutionary questions that are not tractable using molecular evidence, including questions about the evolution of the closest living relatives of modern humans and evolution within our own Hominina clade. In the last few decades the emergence of evolutionary developmental biology has resulted in a resurgence of interest in comparative anatomy, including myology [51,52,69,73-82]. Along with Assis [5], we suggest that the forthcoming decades will see a renaissance in the use of myology in phylogenetic systematics. We hope this review will contribute to this renaissance by stimulating an interest in the use of morphological data in general, and of muscles in particular, for phylogeny reconstruction.
We thank the numerous institutions and individuals that have allowed us to obtain and dissected the primate specimens studied by us, as well as Zuogang Peng, Michel Chardon, Annie Burrows, Sam Dunlap, Ashraf Aziz, Justin Adams and Veronique Barriel for discussions and insightful comments. The contributions of RD was supported by George Washington University (GW) Presidential Merit Fellowship and by a Howard University College of Medicine start-up package, those of LM by Harvard University funds and those of BW by the GW University Professorship in Human Origins and support from the GW Provost via the GW Signature Program.
