Research Article - (2014) Volume 0, Issue 0
The acute toxicity and dose-response studies of methanol extracts of the fruit (MFE), leaf (MLE), bark (MBE) and root (MRE) of Morinda lucida were investigated in adult male albino rats (Wistar strain) weighing between 180 – 250g. They were orally administered (p.o) with the various extracts by observing Lorke’s method. The median lethal dose (LD50) was greater than 5000 mg/kg for all the extracts tested and without any observable sign of toxicity. The genotoxic and geno-protective properties of all the MFE, MLE, MBE and MRE on pUC18 plasmid DNA were also studied, in the presence of Fenton’s reagent using the plasmid nicking assay method. While MFE, MBE and MRE offered better protection to pUC18 plasmid DNA at lower doses particularly at 10 μg/μl, MLE was genotoxic at all the doses as evidenced by the presence of DNA linear forms. While parts of Morinda lucida showed no visible toxicity even at very high doses, they proved to be genotoxic to pUC18 plasmid DNA at higher concentrations.
Keywords: Acute toxicity; Genotoxicity; pUC18 Plasmid DNA; Morinda lucida
Morinda lucida is an important plant in traditional medicine. Decoctions and infusions or plasters of root, bark and leaves are recognized remedies against different types of fever, including yellow fever, malaria, trypanosomiasis, feverish conditions during child birth and even cancer [1,2]. The plant is also employed in cases of diabetes, hypertension, cerebral congestion, dysentery, stomach-ache, ulcers, leprosy and gonorrhea [3]. In Nigeria Morinda lucida is one of the 4 most used traditional medicines against fever.
The administration of an herbal material may invariably have its accompanying hazards. First is the issue of toxicity, mostly due to lack of dosage regimens. Thus, the determination of acute toxicity is the primary step in investigating the toxicity of an unknown substance [4]. Acute toxicity index is referred to as LD50 [5]. Apart from carrying out LD50 investigation for an unknown substance, it is necessary to access the likely toxicity encountered by undiscerning users, and their toxicity limits evaluated [4]. Secondly, during the inevitable process of the biotransformation of xenobiotics, free radicals including reactive oxygen species (ROS) are generated and these are also harmful to the body system.
As a matter of fact, many non-communicable diseases seem to be both interrelated and share the same etiology particularly, with regards to ROS. For instance, evidences are replete that ROS from exogenous sources are involved in the pathogenesis of a large number of diseases including cancer [6,7] diabetes, inflammation [8,9] Alzeimer and Parkinson disease [8] and nucleic acid damage [9,10].
A substantial body of evidence links the production of reactive oxygen species, and subsequently, oxidative stress and damage to the pathogenesis of age-related and chronic diseases [11-13]. Oxidative stress can occur through overproduction of reactive oxygen and nitrogen species through either endogenous or exogenous insults. For instance, genotoxicity which may have commenced with acute inflammation could result in carcinogenesis, and the unregulated or prolonged production of cellular oxidants has been linked to mutation (oxidantinduced DNA damage) as well as modification of gene expression. When the redox balance is shifted in favor of cellular oxidants, oxidative damage to nucleic acids, lipids, or proteins can result and produce modification to cell function and cell viability.
Genotoxicity describes an agent’s damaging ability to the DNA which can cause mutations that can result in cancer. Thus, genotoxicity is a broad term that refers to any deleterious change in the genetic material regardless of the mechanism by which the change is induced [14]. The mechanism for DNA damage, leading to mutation, is due to the attack of hydroxyl radicals on guanine in DNA. Free radicals such as superoxide anion and nitric oxide intervened in the inflammatory process. Nitric oxide can induce cyclo-oxygenase, the rate limiting step enzyme for inflammation process and enhance production of interleukin-1 and tumor necrosis factor [15].
The current increasing global wave of interest in the use of medicinal plants in treating/managing diverse ailments has inadvertently increased the chances of introducing compounds that may be genotoxic or genoprotective into the body. According to [16] studies on the genotoxicity and genoprotective properties of natural plant extracts can help evaluate the safety and effectiveness of herbal products.
Many researchers have probed into the effects of medicinal and herbal plants on human tissue and DNA. It has been revealed that animals possessing highest rates of oxidative damage induced by free radicals generally have the shortest life spans [17]. Thus, the genotoxic or genoprotective assessment of medicinal plant extracts is extremely important because they constitute very attractive sources of compounds that can be used in the development of novel pharmaceutical products [18]. Also, genotoxicity tests form an important part of risk assessment of potential carcinogens [19].
An in vivo assessment of DNA damage and the protective effects of methanolic extracts of Miconia species have indicated a protective effect against cyclophosphamide induced DNA damage [20] and showed the advantages in using crude extracts than the application of isolated compounds in protecting the DNA.
Usually, damage to plasmid DNA results in a cleavage of one of the phosphodiester chains of the supercoiled DNA and produces a relaxed open circular form. Further cleavage near the first breakage results in linear double-stranded DNA molecules. The formation of circular form of DNA is indicative of single-strand breaks and the formation of linear form of DNA is indicative of double-strand breaks [21]. Since the Fenton’s reagents are composed of ascorbic acid, H2O2 and Fe3+, they can produce OH? according to the following reaction [22]
Ascorbic acid + Fe3+ → Ascorbic acid + Fe2+ (1)
Fe2+ + H2O2 + Fe2+ → OH? + OH- (2)
OH- + DNA DNA damage product + OH- → (3)
The formation of Fe2+ after reduction of Fe3+ in the presence of potent reducing agent ascorbic acid, initiate the decomposition of H2O2 leading to generate highly reactive free radical species OH- via Fentonlike reactions above (equations 1 and 2) [9].
Majority of the oxidative damage in the biological system is caused by the OH- [23]. These OH- binds to the DNA resulting to strand breakage, de-oxy sugar fragmentation and base modification [24].
Chemicals/Reagents
Agarose gel electrophoresis kit; Quercetin standard; pUC18 plasmid; Fenton’s reagent.
Equipment
Electrophoresis machine; UV lamp; Camera (JVC Super LoLux).
LD50 determination
The method described by Lorke [5] was employed. This was carried out in two stages using oral route of administration. In stage 1, three different groups of three rats each were administered with different doses of the extracts, namely: 10, 100 and 1000 mg/kg and observed for 24 hours for death or physical signs of toxicity. In stage 2, fresh sets of rats were used. With no death recorded in stage 1, doses of 1000, 1600, 2900 and 5000 mg/kg were administered to four different rats and the animals were observed for another 24 hours for toxicity signs. The median lethal dose (LD50) was calculated based on findings at this stage.
Genotoxicity test: Plasmid nicking assay
Into 0.5 ml micro-centrifuge tubes, 250 ng (0.5 μl) of pUC18 plasmid DNA were added, followed by 10 μl of Fenton’s reagent, then 10 μl of different concentrations [100 μg, 200 μg and 500 μg] of Morinda iextracts or PBS (phosphate buffered saline). The aliquots were raised to a final volume of 20.5 μl then incubated at 37°C for 30 minutes. Three (3 μl) of gel loading dye were added and electrophoresed in a horizontal slab gel apparatus in TAE gel buffer for 45 minutes at 40 V/20 mA. Gels were stained with 0.6 μg/ml of ethidium bromide for 30 minutes followed by de-staining in de-ionized water for another 30 minutes. Photographs of the gels were obtained under UV light and visually assessed. The extent of damages (or otherwise) were observed. Quercetin was used as positive control.
LD50 of M. lucida fruit, leaf, bark and root extracts
The median lethal dose, LD50 of the methanol extracts of the fruit (MFE), leaf (MLE), bark (MBE) and root (MRE) of Morinda lucida were all above 5000 mg/kg. The rats tolerated the extracts even at larger doses with no signs of acute toxicity.
Genotoxic and genoprotective effects of M. lucida fruit, leaf, bark and root extracts on pUC18 plasmid DNA
Figure 1 plates A-D show the plasmid nicking effect of Fenton’s reagent on the pUC18 plasmid DNA (lanes 7) where only the plasmid DNA and Fenton’s reagent were incubated, as no traces of the DNA substances were visible. This contrasted with lanes 8 (pUC18 plasmid DNA+PBS) where the supercoiled pUC18 plasmid DNA was intact and clearly visible, and lanes 3 where the protective effect of quercetin standard (20 μg/μl) was observed.
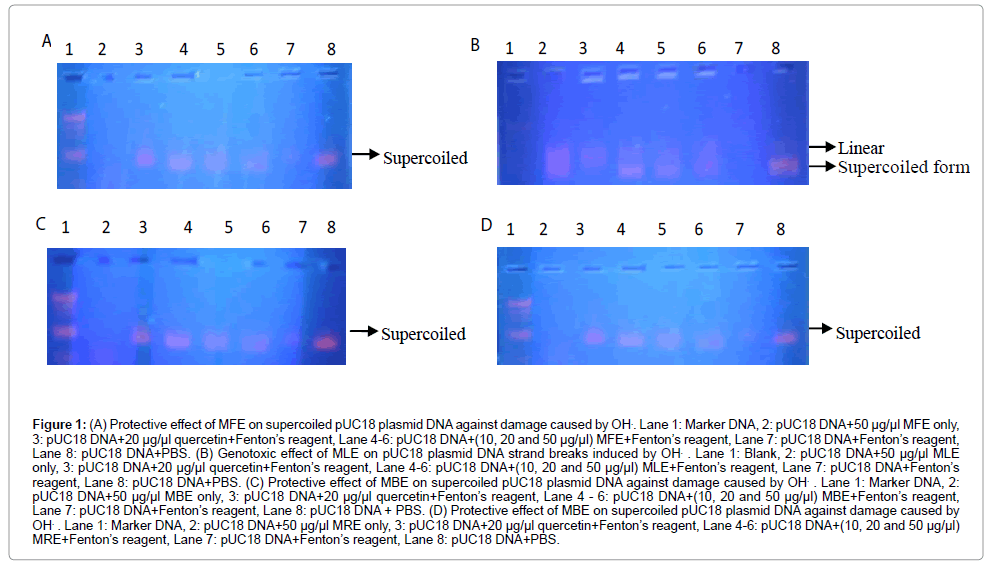
Figure 1: (A) Protective effect of MFE on supercoiled pUC18 plasmid DNA against damage caused by OH.. Lane 1: Marker DNA, 2: pUC18 DNA+50 μg/μl MFE only, 3: pUC18 DNA+20 μg/μl quercetin+Fenton’s reagent, Lane 4-6: pUC18 DNA+(10, 20 and 50 μg/μl) MFE+Fenton’s reagent, Lane 7: pUC18 DNA+Fenton’s reagent, Lane 8: pUC18 DNA+PBS. (B) Genotoxic effect of MLE on pUC18 plasmid DNA strand breaks induced by OH. . Lane 1: Blank, 2: pUC18 DNA+50 μg/μl MLE only, 3: pUC18 DNA+20 μg/μl quercetin+Fenton’s reagent, Lane 4-6: pUC18 DNA+(10, 20 and 50 μg/μl) MLE+Fenton’s reagent, Lane 7: pUC18 DNA+Fenton’s reagent, Lane 8: pUC18 DNA+PBS. (C) Protective effect of MBE on supercoiled pUC18 plasmid DNA against damage caused by OH. . Lane 1: Marker DNA, 2: pUC18 DNA+50 μg/μl MBE only, 3: pUC18 DNA+20 μg/μl quercetin+Fenton’s reagent, Lane 4 - 6: pUC18 DNA+(10, 20 and 50 μg/μl) MBE+Fenton’s reagent, Lane 7: pUC18 DNA+Fenton’s reagent, Lane 8: pUC18 DNA + PBS. (D) Protective effect of MBE on supercoiled pUC18 plasmid DNA against damage caused by OH. . Lane 1: Marker DNA, 2: pUC18 DNA+50 μg/μl MRE only, 3: pUC18 DNA+20 μg/μl quercetin+Fenton’s reagent, Lane 4-6: pUC18 DNA+(10, 20 and 50 μg/μl) MRE+Fenton’s reagent, Lane 7: pUC18 DNA+Fenton’s reagent, Lane 8: pUC18 DNA+PBS.
There was no visible band in lane 2 where only the pUC18 plasmid DNA was incubated with 50 μg/μl of MFE, MBE and MRE, depicting the genotoxic effect of the extracts of fruit, bark and root of M. lucida at the highest concentration used (50 μg/μl). Lane 1 served as blank (plate B) or DNA marker (ladder) made of varying kilobytes of DNA (plates A, C and D).
Figure 1 lanes 4 to 6 of plates A, C and D, showed the protective effect of the MFE, MBE and MRE on the pUC18 DNA, where the pUC18 DNA were incubated with 10 μl of Fenton’s reagent plus 10, 20 and 50 μg/μl of each of MFE, MBE and MRE respectively. The intensities of the supercoiled bands were more visible at lower concentrations of each of the extracts. The highest visibilities of the supercoil were observed in lanes 4 where 10 μg/μl of the various extracts was used.
The MLE on the other hand, showed a different activity, specifically, genotoxicity in all its presentations. As shown in Figure 1 lanes 4 to 6 of plate B, there was absence of protective effect of the MLE on pUC18 DNA, where the pUC18 DNA was incubated with 10 μl of Fenton’s reagent plus 10, 20 and 50 μg/μl of the MLE respectively. However, the intensity of the supercoiled band was relatively more visible with lower concentrations (Plate 5). The highest visibility of the supercoil was observed in lane D where 10 μg/μl of the methanolic leaf extract of M. lucida was used.
The result of acute oral toxicity study in the albino wistar rats recorded an LD50 of the methanolic extracts of all the parts of M.lucida plant as being above 5000 mg/kg, as the experimental rats tolerated the extracts without any symptoms of acute toxicity (no mortality, skin changes, aggressiveness, diarrhoea, restiveness, seizures, dizziness, weakness, or withdrawal from either food or water) even at larger doses of extracts administered. Lower LD50 implies a more toxic extract being tested. According to Lorke [5], an LD50 value beyond 5000 mg/kg is of no toxicological significance
Morinda lucida has been used in managing some ailments and known as a very good anti-oxidant/scavenger of reactive metabolites [25]. Similarly, polyphenols-rich red onion peel was observed to have offered protection to pBR322 plasmid DNA against the damage caused by Fenton’s reagent by a mechanism of electron or hydrogen-atom transfer [26].
In this work however, it was observed that the methanol leaf extract offered least protection to the pUC18 plasmid DNA against the damage caused by Fenton’s reagent. The loss of DNA protective abilities seemed to have increased with increasing concentrations of the leaf extract, evidenced by the presence of linear forms in lanes 4 to 6 of plate B and the fading intensities observed as one moves from lane 4 to 6. It was also observed that there seemed to have been a combined damaging effect of the extract and the Fenton’s reagent because, while the plasmids were incubated with the same amount of Fenton’s reagent, they were treated with different concentrations of MLE. In that, lane 6 with the highest extract concentration (50 μg/μl) showed a very faint or an almost non-visible band while lane 4 with the least concentration (10 μg/μl) showed the most visible band. The reason for the faint or non-visibility of DNA bands are because, to make the DNA visible in the gel, ethidium bromide is added as the staining agent which binds to DNA by inserting itself between the base-pairs by “intercalation” which can then be seen by exposure of the gel to UV-light, due to the increased fluorescence upon binding to the DNA. But small fragment are hardly visible [27](SCL, 2014). Also, though ethidium bromide can be used to detect both single- and double-stranded nucleic acids, the affinity of the dye for single-stranded nucleic acid is relatively low and the fluorescent yield is comparatively poor [21].
A different scenario from the MLE was observed with MFE, MBE and MRE [plates A, C and D, respectively]. While every materials and conditions were the same, there was no genotoxic effect observed with MFE, MBE and MRE, evidenced by the absence of the linear strand breaks and/or, the nicked circular forms. However, it was observed that the bands became less visible with increasing concentrations of the extracts.
Observed also was lanes 2 (plates A, C and D) where the highest dose (50 μg/μl) of only the extracts were added to the pUC18 plasmid DNA (without Fenton’s reagent), where there were no visible bands; an evidence that the entire DNA was damaged in each case, similar to the event that may have taken place in lanes 7 where pUC18 plasmid DNA and Fenton’s reagent only were incubated.
It may thus be reasoned that the MFE, MBE and MRE may have conferred protection against the damaging effect of Fenton’s reagent at lower concentrations but genotoxic at higher doses. Furthermore, the DNA protective capabilities of MFE, MBE and MRE at the lowest dose of 10 μg/μl (lanes 4) showed protection, similar to the quercetin standard (lanes 3) at concentration of 20 μg/μl. Parts of M. lucida have been acknowleged as excellent antioxidants and good scavengers of reactive metabolites such as OH?, O2?-, and ROO? [25,29].
In this experiment, it was possible to examine the oxidative DNA damage or the protective properties of the fruit, leaf, bark, and root extracts against the OH?-induced damage on pUC18 plasmid DNA. As observed in Figure 1, the methanol leaf extract of the M. lucida was unable to protect the pUC18 plasmid DNA from oxidative damage
In a related experiment, [9] showed that after incubation of pUC18 plasmid DNA with Fenton’s reagent for 45 min, there was a resultant cleavage of the supercoil to yield an open circular and linear forms of plasmid DNA, indicating that OH? generated from iron-mediated decomposition of H2O2 produced both single-strand and double-strand DNA breaks [30], reported that H2O2 and ?O2- are potentially cytotoxic; most of the oxidative damage in biological systems is caused by the OH?, which is generated by the reaction between H2O2 and ?O2- in the presence of redox-active metals. This type of oxidative damage can be reduced in the presence of standard antioxidants, which converts H2O2 into water and oxygen. In the present experiment, inclusion of the fruit, bark and root extracts at the lowest concentration of (10 μg/μl) to DNA and Fenton’s reagent mixture induced the significant reduction in the formation of open circular and linear forms and increased supercoiled or native form of plasmid DNA.
Also, [9] showed that the aqueous fruit and seed extracts of Moringa oleifera at concentrations of between (10-25 μg/ml) and trolox (10-25 μM), showed partial and complete protection of supercoiled DNA, and mitigated the oxidative stress produced by Fenton-like reactionmediated OH?. Recently, studies have shown that redox-active metals in solution could bind to phenolic antioxidant compounds and form complexes, and consequently prevent the reduction of redox-active metal ions with H2O2 [22,31].
The M. lucida methanol fruit, bark and root extracts used in this experiment, like the Moringa oleifera extracts investigated by Singh et al. [4] might prevent the reaction ofFe3+ with H2O2. On the other hand, natural polyphenols can influence the free radicals mediated oxidation of DNA through simple mechanism including quenching of reactive oxygen species by donating hydrogen atom or electron. This type of action might diminish the reduction potential ofFe3+ leading to the inhibition of Fenton-like reaction, and might also directly scavenge OH? and therefore protecting the supercoiled plasmid DNA from OH?- dependent strand breaks.
