Research Article - (2014) Volume 2, Issue 5
Water shield (Brasenia schreberi J. F. Gmel) is an aquatic plant cultivated in Northern Japan, from which we have isolated polyphenolic compounds including a novel flavonol glycoside, junsainoside A. Recently, dermal fibroblasts have reported to be affected by adipocytokines from subcutaneous adipocytes. Hence, we evaluated the effects of polyphenolic compounds from water shield on fat accumulation, adipocytokine release and adipocytokine-induced fibroblast status. In differentiated 3T3-L1 adipocytes, ethyl gallate, caffeoyl glucose, hypolaetin 7-O-glucoside, kaempferol and junsainoside A significantly suppressed lipid accumulation at 10 μM. In addition to these constituents, ferulic acid, kaempferol 3-O-(6"-galloyl) glucoside, quercetin 3-O-(6"-galloyl) glucoside, quercetin 3-O-glucoside and gossypetin suppressed the lipid accumulation in human subcutaneous adipocytes. Ethyl gallate and junsainoside A also enhanced adiponectin production from subcutaneous adipocytes without induction of TNF-α release. On the other hand, ethyl gallate and quercetin recovered from TNF- α-induced fibroblast damage. Moreover, ferulic acid, quercetin and kaempferol enhanced the expression of hyaluronan binding protein 2 and hyaluronan synthase 2 in adiponectintreated fibroblasts. These results suggested that water shield contains various types of polyphenolic compounds with anti-adipogenic activity, some of which protect dermal fibroblasts from TNF-α damage and probably enhance hyaluronan production. It is assumed that polyphenols in water shield maybe suppress subcutaneous fat accumulation and maintain dermal fibroblast and hyaluronan-related status.
Keywords: Water shield; Junsainoside A; Adipocyte; TNF- α; Adiponectin; Hyaluronan
TNF: Tumor Necrosis Factor-α,β; WSE: Water shield extract, DEPT: Distortionless enhancement by polarization transfer, HMQC: Heteronuclear multiple quantum coherence spectroscopy, HMBC: Heteronuclear multiple bond connectivity, RIPA : Radioimmunoprecipitation assay, HAS: Hyaluronan synthase, HABP: Hyaluronan binding protein, HRP: Horseradish peroxidase , D-MEM: Dulbecco’s modified essential medium, FCS: Fetal calf serum, DMSO: Dimethyl sulfoxide, MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide.
Water shield (Brasenia schreberi J. F. Gmel, Cabombaceae) is an aquatic plant that grows in Asia, Australia, Africa and North America. As the surfaces of its leaves and buds are covered with mucus consisting of polysaccharide [1], the texture is similar to gelatine. In Japan, water shield (junsai in Japanese) is cultivated as a vegetable and the young buds are used as a food ingredient. On the other hand, the leaves have been used as stomachic in China; however, the constituents of water shield have not been well studied. Only quercetin 7-O-glucoside and gallic acid have been reported as principal polyphenols [2]. In terms of the biological activity of the extract from water shield, anti-oxidative [2], anti-inflammatory [2] and hypolipidemic activities [3] have been reported. Considering this background, we performed isolation of the constituents of water shield leaves and identified various types of polyphenol including a new flavonol glycoside named junsainoside.
A number of polyphenols have been reported to suppress visceral fat accumulation in adipocytes. 3T3-L1 preadipocytes [4] and primary cultured visceral adipocytes [5] have been frequently used for the assay. These results indicate the efficacy of polyphenols against obesityrelated insulin resistance, dyslipidemia and arteriosclerosis. However, from the perspective of dermatology, the effect of polyphenols on subcutaneous fat accumulation has not been well studied. Recently, the relationship between subcutaneous adipocytes and dermal fibroblasts has been focused on. Subcutaneous fat exists in the hypodermis and releases adipocytokines including tumor necrosis factor (TNF) - α and adiponectin [6]. TNF- α is also released from dermal inflammatory cells and induces fibroblast apoptosis [7]. On the other hand, adiponectin enhances hyaluronan and collagen production from fibroblasts [8]. Thus, TNF- α and adiponectin affect dermal fibroblasts with negative and positive responses and the balance of these adipocytokines is considered to be crucial to maintain stable dermal conditions. Against the above background, we investigated the effect of water shield extract (WSE) and its constituents on lipid accumulation and adipocytokine release in subcutaneous adipocytes. In addition, the effects of the constituents on fibroblasts treated with adipocytokines were evaluated.
Preparation of WSE
The leaves of water shield cultivated in Mitane town (Akita prefecture, Japan) were harvested in June 2011. The leaves were airdried and ground. The powdered leaves (200 g) were then extracted with 30 w/w% ethanol (EtOH, 6 kg) at 50°C for 2 h. The solvent was evaporated to obtain WSE (44.3 g, yield: 22.1%). The polyphenol content determined by the Folin-Ciocalteu method was 27.3% (gallic acid equivalent).
Isolation of the compounds in water shield
Dried powder of water shield (870 g) was extracted with 50 w/w% EtOH (20 kg) at 80°C for 3 h. The solvent was evaporated to obtain 50% EtOH extract (188 g, yield: 21.6%). The extract (100 g) was applied to Diaion® HP20 (Mitsubishi Chemical Corporation, Tokyo, Japan) column chromatography (1 kg). The column was eluted with 10, 30, 70 and 100 v/v% methanol (MeOH) sequentially to obtain a 10% MeOH fraction (2.3 g), 30% MeOH fraction (14.2 g), 70% MeOH fraction (29 g) and 100% MeOH fraction (4.7 g). The 10% MeOH fraction (2.3 g) was applied to octadecyl silyl (ODS, Fuji Silysia Chemical Ltd., Kasugai, Japan) silica gel column chromatography (50 g, 10→20→30% MeOH) to obtain fractions: fraction (Fr.) 1 (0.79 g), Fr. 2 (0.75 g) and Fr. 3 (0.42 g). Fr. 2 (0.75 g) was purified by HPLC (C30 column (Develosil RPAQUEOUS; Nomura Chemical Co. Ltd., Seto, Japan, 250×20 mm i.d.), 10% MeOH) to obtain phenylalanine (1, 2 mg, yield: 0.002% from 50% EtOH extract). Fr. 3 (0.42 g) was separated by HPLC (C30 column, 50% MeOH) to obtain p-phenylbenzoic acid (2, 2 mg, 0.002%) [9]. The 70% MeOH fraction (29 g) was applied to ODS column chromatography (300 g, 40→50→60→70→100% MeOH) to obtain fractions: Fr. 1 (23.9 g), Fr. 2 (1.4 g), Fr. 3 (1.8 g), Fr. 4 (0.4 g) and Fr. 5 (0.2 g). Fr. 1 (1 g) was purified by HPLC [ODS column (Inertsil preparative ODS; GL Science, Tokyo, Japan, 250×20 mm i.d., 40% MeOH) and C30 column (60% MeOH)] to obtain ethyl gallate (3, 3.2 mg, 0.076%) [10], ferulic acid (4, 0.6 mg, 0.014%) and caffeoyl glucose (5, 1 mg, 0.024%) [11]. Fr. 2 (1.4 g) was purified by HPLC (ODS column (50% MeOH) and C30 column (50-60% MeOH)) to obtain hypolaetin 7-O-glucoside (6, 1 mg, 0.001%) [12], kaempferol 3-O-(6"-galloyl) glucoside (7, 6 mg, 0.006%) [13], quercetin 3-O-(6"-galloyl) glucoside (8, 8 mg, 0.008%) [14], quercetin 7-O-glucoside (9, 2 mg, 0.002%) [15], quercetin 3-O-glucoside (10, 26 mg, 0.026%) [16] and gossypetin (11, 7 mg, 0.007%) [17]. Fr. 3 (1.8 g) was purified by HPLC (ODS column (60% MeOH) and C30 column (65- 70% MeOH)) to obtain junsainoside A (12, 32 mg, 0.032%), quercetin (13, 62 mg, 0.062%) and kaempferol 7-O-(6"-galloyl) glucoside (14, 2 mg, 0.002%) [18]. The 100% MeOH eluent (4.65 g) was applied to ODS column chromatography (100 g, 70→80→90→100% MeOH) to obtain fractions: Fr. 1 (1.62 g), Fr. 2 (0.77 g), Fr. 3 (0.67 g) and Fr. 4 (0.95 g). Fr. 1 (1.62 g) was purified by HPLC (ODS column (70-80% MeOH) and C30 column (70-80% MeOH)) to obtain quercetin (13, 84 mg, 0.084%) and kaempferol (15, 54 mg, 0.055%). A refractive index detector was used for detection of the peak on HPLC purification. Isolated known compounds were identified by comparison of 13C- and 1H-NMR spectra and mass spectra with reported values or authentic standard samples of 1, 4, 13 and 15 (Figure 1).
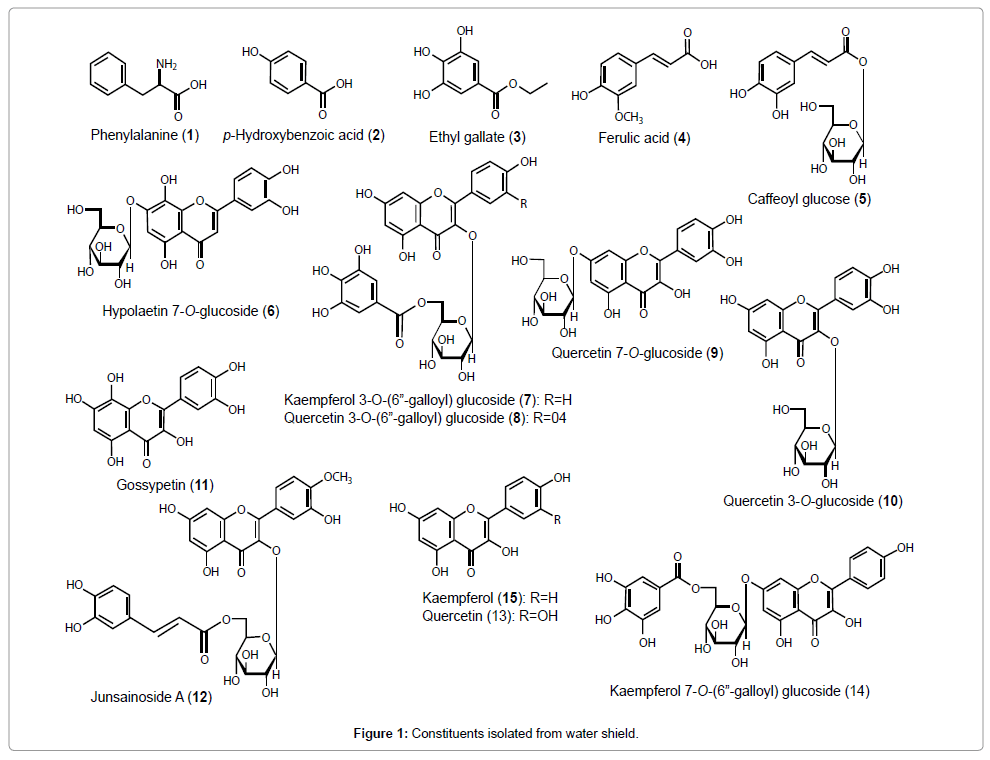
Figure 1: Constituents isolated from water shield.
Structural elucidation of junsainoside A (12)
Junsainoside A (12) was isolated as a pale yellow powder with negative optical rotation ([α]D21 –52.0° in MeOH). The IR spectrum of 12 suggested the presence of hydroxyl (3400 cm–1), α, β-unsaturated ester (1689 cm–1), γ-pyrone (1655 cm–1), aromatic ring (1605, 1514 cm–1) and ether (1016 cm–1). In the positive-ion matrix-assisted laser desorption/ionization (MALDI)-MS of 12, a quasimolecular ion peak was observed at m/z 663 (M+Na)+, and high-resolution (HR) MALDIMS analysis of the quasimolecular ion peak revealed the molecular formula of 12 to be C31H28O15. Acid hydrolysis of 12 with 5% aqueous H2SO4-1,4-dioxane yielded D-glucose, which was identified by HPLC of the tolylthiocarbamoyl thiazolidine derivative. To determine the absolute configuration of glucose of 12, the method reported by Tanaka et al. was used [19]. The 1H-NMR (CD3OD) and 13C-NMR spectra of 12, which were assigned by various NMR experiments, showed signals assignable to a tamarixetin part [δ 6.07 (1H, d, J=2.0 Hz, H-6), 6.25 (1H, d, J=2.0 Hz, H-8), 6.80 (1H, d, J=8.8 Hz, H-5'), 7.56 (1H, dd, J=8.8, 2.0 Hz, H-6'), 7.57 (1H, d, J=2.0 Hz, H-2')], a β-D-glucopyranosyl moiety [δ 4.28 (1H, m, 6''-H2), 3.35 (1H, m, 4-H), 3.45 (1H, m, 5-H), 3.43 (1H, m, 3-H), 3.50 (1H, m, 2-H), 5.26 (1H, d, J=7.3 Hz, 1''-H)], and a (E)-caffeoyl group [δ 6.12 (1H, d, J=16.0 Hz, 8'''-H), 6.79 (1H, d, J=8.3 Hz, 3'''-H), 6.91 (1H, dd, J=2.0, 8.3 Hz, 2'''-H), 7.04 (1H, d, J=2.0 Hz, 6'''-H), 7.39 (1H, d, J=16.0 Hz, 7'''-H)]. The 1H and 13C NMR spectra of 12 were assigned with the aid of distortionless enhancement by polarization transfer (DEPT), heteronuclear multiple quantum coherence spectroscopy (HMQC) and heteronuclear multiple bond connectivity (HMBC) experiments. The postions of a (E)-caffeoyl group and a β-D-glucopyranosyl moiety were confirmed by HMBC spectroscopy (Figure 2). Namely, long-range correlations were observed between the following proton and carbon pairs: 1''-H and 3-C; 6''-H and 9'''-C. Comparison of the 13C-NMR data for tamarixetin 3-O- β -D-glucopyranoside [20] with those for 12 revealed an acylation shift around the 6'' position. From this evidence, the chemical structure of junsainoside A was determined to be tamarixetin 3-O-(6''-O-Ecaffeoyl)- β -D-glucopyranoside (12).
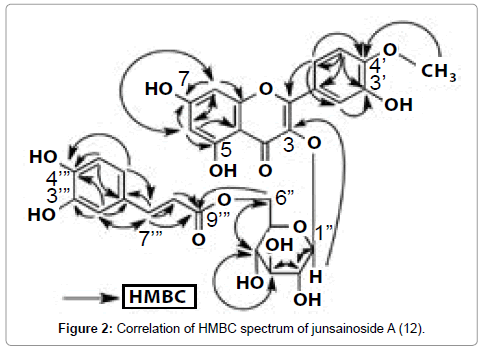
Figure 2: Correlation of HMBC spectrum of junsainoside A (12).
Junsainoside A (12): A pale yellow powder, [α]D21 –52.0° (c=0.1, MeOH). HR-MALDI-TOF-MS Calcd for C31H28O15Na (M+Na)+: 663.1320; Found: 663.1327, UV [MeOH, nm, (log ε)]: 255 (4.28), 329 (4.32). IR (KBr, cm–1): 3400, 1689, 1655, 1605, 1514, and 1016. 1H-NMR (CD3OD, 500 MHz) δ: 3.35 (1H, m, 4''-H), 3.43 (1H, m, 3''-H), 3.45 (1H, m, 5''-H), 3.50 (1H, m, 2''-H), 3.88 (3H, s, 4'-OCH3), 4.28 (2H, m, 6''-H), 5.26 (1H, d, J=7.3 Hz, 1''-H), 6.07 (1H, d, J=2.0 Hz, 6-H), 6.12 (1H, d, J=16.0 Hz, 8'''-H), 6.25 (1H, d, J=2.0, 8-H), 6.79 (1H, d, J=8.3 Hz, 5'''-H), 6.80 (1H, d, J=8.8 Hz, 5'-H), 6.91 (1H, dd, J=8.3, 2.0 Hz, 6'''- H), 7.04 (1H, d, J=2.0 Hz, 2'''-H), 7.39 (1H, d, J=16.0 Hz, 7'''-H), 7.56 (1H, dd, J=8.8, 2.0 Hz, 6'-H), 7.57 (1H, d, J=2.0 Hz, 2'-H). 13C-NMR (CD3OD, 125 MHz) δ c: 159.2 (C-2), 135.2 (C-3), 179.4 (C-4), 162.9 (C-5), 99.8 (C-6), 165.8 (C-7), 94.7 (C-8), 158.4 (C-9), 105.6 (C-10), 123.1 (C-1'), 123.4 (C-2'), 115.9 (C-3'), 149.8 (C-4'), 115.9 (C-5'), 123.4 (C-6'), 56.5 (4'-OCH3), 103.8 (Glc-1''), 75.6 (Glc-2''), 78.0 (Glc-3''), 71.8 (Glc-4''), 75.9 (Glc-5''), 64.3 (Glc-6''), 127.6 (C-1"'), 146.8 (C-2"'), 149.3 (C-3"'), 150.5 (C-4"'), 116.4 (C-5"'), 124.2 (C-6"'), 146.8 (C-7"'), 115.0 (C-8"'), 168.8 (C-9"'). Positive-ion MALDI-TOF-MS: m/z 663 (M+Na)+ (Figure 2).
Reagents
Human TNF-α quantkine ELISA kit, human total adiponectin/ Acrp30 quantkine ELISA Kit and recombinant human adiponectin/ Acrp30 were obtained from R&D Systems (Minneapolis, MN, USA). Recombinant TNF-α was purchased from Life Technologies (MD, USA). Kaempferol 3-O-glucoside and tiliroside were purchased from Extrasynthese (Cedex, France). Radioimmunoprecipitation assay (RIPA) buffer, protease and phosphatase inhibitor cocktail, and Pierce Western Blotting Substrate Plus were purchased from Thermo Fisher Scientific Inc. (Waltham, MA, USA). Goat anti-hyaluronan synthase (HAS) 2 IgG was obtained from Santa Cruz Biotechnology (Dallas, TX, USA). Rabbit anti-hyaluronan binding protein (HABP) 2 IgG was purchased from Abnova (Taipei, Taiwan). Mouse anti-β-actin (H- 196) IgG and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Horseradish peroxidase (HRP)-conjugated anti-rabbit IgG, antigoat IgG and anti-mouse IgG were purchased from Millipore (Billerica, MA, USA).
Cells
3T3-L1 (JCRB9014) and human diploid fibroblasts (TIG-108, JCRB0537) were obtained from the National Institute of Biomedical Innovation (Osaka, Japan). Normal human neonatal fibroblasts (NB1RGB, RCB0222) were obtained from Riken Bio Resources Center (Tsukuba, Japan). Human subcutaneous preadipocyte culture kit (Poietics™) was obtained from Lonza (Basel, Switzerland).
Lipid accumulation in cultured 3T3-L1 and human subcutaneous adipocytes
3T3-L1 cells (1×104 cells/200 μL) suspended in Dulbecco’s modified essential medium (D-MEM) containing 10% fetal calf serum (FCS) were seeded onto a 48-well culture plate and cultured for 24 hr. The medium was then changed to a new one containing 10% FCS, 5 μg/mL insulin, 0.25 μM dexamethasone and 0.5 mM isobutylmethyl xanthine and the cells were cultured for 2 days to induce differentiation. The medium was then changed to a new one containing 10% FCS, 5 μg/ mL insulin and test sample dissolved in dimethyl sulfoxide (DMSO). The final concentration of DMSO was 0.1%. The medium was changed to a new one every other day and the cells were cultured for a total of 6 days. The cells were stained with oil red O and observed under a microscope. The dye in cells was dissolved in isopropanol to measure the absorption at 540 nm. For the evaluation of cytotoxicities of the compounds from water shield, MTT assay was performed according to Denizot et al. [21], with modification. Briefly, cells (5×103 cells/100 μL) were seeded in a 96-well culture plate and treated by the same procedure as described above. Then MTT reagent was added to each well and the plate was incubated for 2-4 hr. After washing the plate with phosphate buffered saline, pH 7.4 (PBS, Gibco®, Life Technologies, CA), the formed formazan dye was dissolved in 0.04M HCl/isopropanol solvent completely and the absorbance was measured at 570 nm.
Human subcutaneous preadipocytes (2×104 cells/200 μL) suspended in the growth medium attached to the kit were seeded onto a 48-well culture plate and cultured for 24 hr. The medium was changed to the differentiation medium containing test sample (final concentration of DMSO: 0.1%) for 10 days. The medium was then collected and stored at -20ºC. The contents of TNF-α and adiponectin were determined using a commercial kit. The cells were stained with oil red O and absorbance was measured as described above.
Fibroblast damage induced by TNF-α
NB1RGB cells (5×103 cells/100 μL) suspended in D-MEM containing 10% FCS were seeded onto a 96-well culture plate and cultured for 24 hr. Then, sample (final concentration of DMSO: 0.1%) and TNF- α (10 ng/mL) were added and cultured for 20 hr. The cytotoxicity by TNF- α was evaluated by MTT assay as described above.
Evaluation of HAS2 and HABP2 expression in fibroblasts induced by adiponectin
TIG-108 cells (1×105 cells/2 mL) suspended in D-MEM containing 10% FCS were seeded onto a 6-well culture plate and cultured for 24 hr. The medium was changed to D-MEM containing 0.5% FCS and cultured for 24 hr. Then, sample (final concentration of DMSO: 0.1%) and adiponectin (1 ng/mL) were added and cultured for 72 hr. Cells were collected with 200 μL of RIPA buffer containing protease and phosphatase inhibitor cocktail. The protein concentration in the cell lysate was adjusted with RIPA buffer to 0.5 mg/mL. The lysate was mixed with the same volume of Laemmli sample buffer (62.5 mM Tris-HCl, 2% SDS, 5% 2-mercaptoethanol, 25% glycerol and 0.01% bromophenol blue) and heated at 95ºC for 5 min. The sample solution (15 μL) was electrophoresed on 10% SDS gel. Separated protein was then transferred to a polyvinylidene fluoride membrane. The membrane was treated with primary antibodies at the following dilution ratios: anti- HAS2 (1: 1,000), anti-HABP2 (1: 2,000) and anti-β-actin (1: 10,000) for 1 hr. HRP-conjugated anti-goat IgG (1:25,000) or anti-rabbit IgG (1: 25,000) was used as the secondary antibody. Detection was performed using by Pierce Western Blotting Substrate Plus and an imaging system (Image Quant LAS500, GE Health Care, Fairfield, CT, USA).
Statistics
The results are expressed as means and S.E. Significance of the differences was examined by one-way ANOVA followed by Dunnett’s test. Differences of p<0.05 were considered significant.
Effects of WSE and the constituents of water shield on lipid accumulation in 3T3-L1 and human cultured subcutaneous adipocytes
As shown in Figure 3, WSE and its constituents showed no cytotoxicity to 3T3-L1 adipocytes. Figures 4A and 4B illustrate lipid accumulation in 3T3L1 adipocytes. WSE (1, 10 μg/mL) significantly suppressed lipid accumulation in the cells. WSE also suppressed lipid accumulation in human subcutaneous adipocytes (Figures 4C and 4D). We also examined the effect of the constituents of water shield extract on lipid accumulation in both cultured cells. Ethyl gallate (3), caffeoyl glucose (5), hypolaetin 7-O-glucoside (6), junsainoside A (12) and kaempferol (15) significantly suppressed lipid accumulation at 10 μM in 3T3-L1 (Table 1). Ethyl gallate (3), 6 and 12 exhibited suppressive activity of more than 50% at 10 μM. In human subcutaneous adipocytes, 3, ferulic acid (4), 5, 6, quercetin 3-O-(6"-galloyl) glucoside (8), quercetin 3-O-glucoside (10), junsainoside A (12) and kaempferol suppressed lipid accumulation at 1 μM. Kaempferol 3-O-(6"-galloyl) glucoside (7) and gossypetin (11) significantly suppressed the accumulation at 10 μM. Among the constituents, 4, 6, 12 and 15 exhibited a suppressive effect of more than 40% at 10 μM.
Figure 3: Cytotoxicity of WSE and its constituents in 3T3-L1 adipocytes.
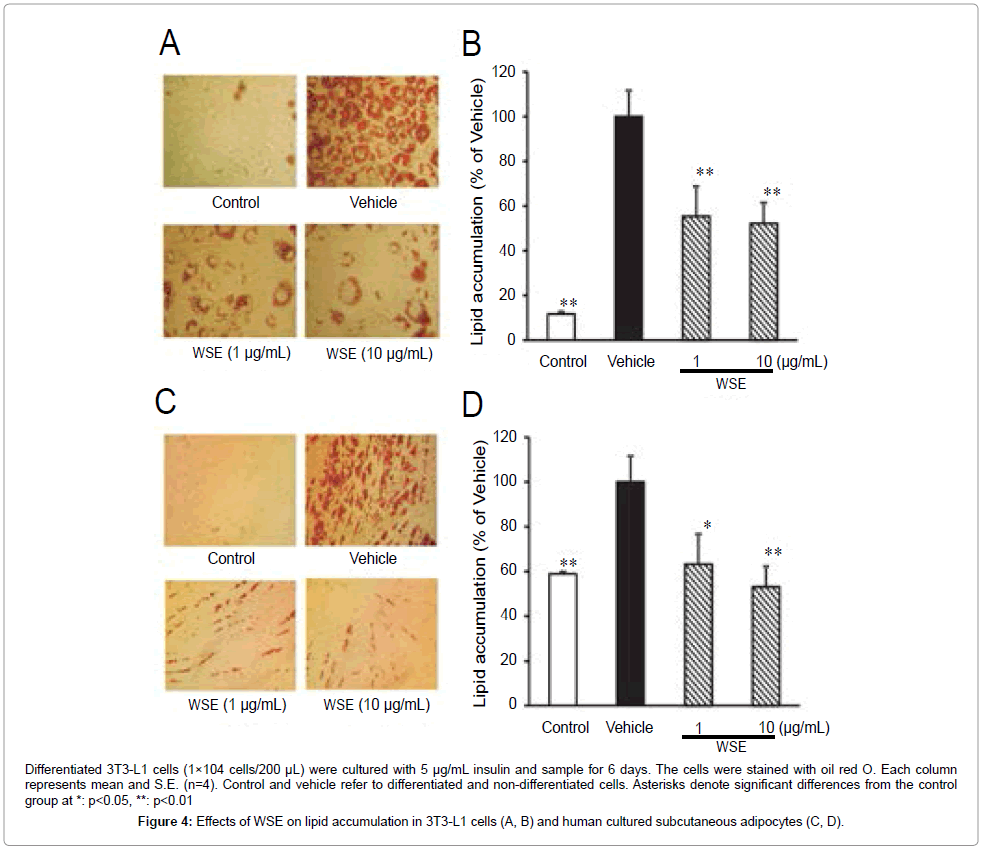
Figure 4: Effects of WSE on lipid accumulation in 3T3-L1 cells (A, B) and human cultured subcutaneous adipocytes (C, D).
| Inhibition (%) | ||||
|---|---|---|---|---|
| 3T3-L1 | Human Subcutaneous Adipocytes | |||
| 1 (μM) | 10 | 1 (μM) | 10 | |
| Ethyl gallate (3) | 63.2±13.9* | 58.9±14.8* | 19.7±4.7** | 34.9±1.9** |
| Ferulic acid (4) | – | – | 23.3±3.1** | 48.5±0.4** |
| Caffeoyl glucose (5) | 42.3±15.3 | 43.6±13.4* | 27.5±1.6** | 35.9±3.1** |
| Hypolaetin 7-O-glucoside (6) | 52.0±17.3* | 55.8±14.1* | 33.5±5.4** | 42.5±1.6** |
| Kaempferol 3-O-(6"-galloyl) glucoside (7) | 25.8±11.9 | 34.6±6.8 | 13.5±4.9 | 25.5±3.8** |
| Quercetin 3-O-(6"-galloyl) glucoside (8) | 35.8±18.5 | 10.8±15.2 | 15.4±3.7* | 21.3±4.5** |
| Quercetin 3-O-glucoside (10) | 2.6±21.6 | 12.8±16.6 | 17.4±3.9* | 22.0±2.6** |
| Gossypetin (11) | 35.9±7.6 | 38.5±11.3 | 12.6±4.3 | 28.4±4.3** |
| Junsainoside A (12) | 44.2±8.0** | 52.8±9.0** | 36.6±6.2** | 45.5±2.4** |
| Quercetin(13) | 30.7±17.7 | 31.1±1.5 | – | – |
| Kaempferol (15) | 35.6±7.5** | 39.9±5.6** | 31.4±4.4** | 40.3±2.4** |
Table 1: Effects of the Constituents Isolated from Water Shield on Lipid Accumulation in Differentiated 3T3-L1 and Human Subcutaneous Adipocytes.
Effects of WSE and the constituents of water shield on TNF-α and adiponectin production from human cultured subcutaneous adipocytes
We evaluated the effect of WSE on TNF- α and adiponectin production from subcutaneous adipocytes. TNF- α production was not suppressed by the treatment of WSE or the constituents (Table 2). On the other hand, WSE did not enhance adiponectin production; however, ethyl gallate (3) and junsainoside A (12) significantly enhanced adiponectin production at 10 μM.
| Amount of adipocytokine production (% of control) | ||||
|---|---|---|---|---|
| TNF-α | Adiponectin | |||
| 1 (µg/mL) | 10 | 1 | 10 | |
| WSE | 110.6±2.2 | 93.2±3.0 | 118.2±8.9 | 111.7±5.1 |
| 1 (µM) | 10 | 1 | 10 | |
| Ethyl gallate (3) | 89.9±5.34 | 86.7±4.5 | 122.2±5.8 | 138.3±10.0** |
| Ferulic acid (4) | 97.7±3.1 | 83.8±6.1 | 120.6±8.5 | 123.3±7.6 |
| Caffeoyl glucose (5) | 112.0±10.4 | 103.6±2.4 | 105.7±5.2 | 107.6±3.7 |
| Hypolaetin 7-O-glucoside (6) | 116.5±6.1 | 109.3±4.7 | 120.6±11.3 | 104.1±4.1 |
| Junsainoside A (12) | 111.0±4.9 | 123.0±12.1 | 130.1±7.9 | 154.5±24.7* |
| Kaempferol (15) | 97.0±3.3 | 92.7±3.2 | – | – |
Table 2: Effects of the Constituents Isolated from Water Shield on TNF-α and Adiponectin Production from Human Subcutaneous Adipocytes.
Effects of WSE and the constituents of water shield on fibroblast damage induced by TNF-α
By the treatment of 10 ng/mL TNF- α, the formazan production indicating cell activity was reduced by approximately 41% compared with that of cells without TNF-α. In WSE-treated cells, the cell activity was significantly recovered from cell damage at 1 and 10 μg/mL. Among the constituents of water shield, ethyl gallate (3) and quercetin (13) significantly recovered the cell activity at 10 μM (Table 3).
| MTT assay(% of control) | ||
|---|---|---|
| Normal | 141.8±13.2* | |
| Control (with TNF-α) | 100.0±8.2 | |
| 1 (μg/mL) | 10 | |
| WSE | 133.1±12.6* | 161.6±11.3** |
| 1 (μM) | 10 | |
| Ethyl gallate (3) | 129.8±6.0 | 135.1±4.6* |
| Ferulic acid (4) | 104.9±8.1 | 84.3±5.4 |
| Caffeoyl glucose (5) | 112.6±3.3 | 129.1±8.6 |
| Hypolaetin 7-O-glucoside (6) | 120.4±14.2 | 98.2±6.2 |
| Kaempferol 3-O-(6"-galloyl)-glucoside (7) | 126.9±4.5 | 117.2±8.2 |
| Quercetin 3-O-(6"-galloyl)-glucoside (8) | 106.2±6.2 | 110.6±8.0 |
| Quercetin 3-O-glucoside (10) | 105.3±6.2 | 124.8±19.5 |
| Gossypetin (11) | 95.5±3.7 | 122.4±9.7 |
| Junsainoside A (12) | 97.8±2.7 | 92.3±7.7 |
| Quercetin (13) | 111.5±7.1 | 150.0±12.6* |
| Kaempferol(15) | 74.4±3.0* | 90.1±2.0 |
Table 3: Effects of WSE and the Constituents Isolated from Water Shield on Fibroblast Damage Induced by TNF-α.
Effects of WSE and the constituents of water shield on the expression of HABP2 and HAS2 in fibroblasts treated with adiponectin
In the fibroblasts cultured with adiponectin, WSE (10 μg/mL) enhanced HABP2 and HAS2 expression (Figure 5). Among the constituents in water shield, ferulic acid (4), quercetin (12) and kaempferol (15) enhanced the expression of HABP2 and HAS2 at 10 μM. Glycosides (3, 5, 6, 7, 8, 10, 12) and gossypetin (11) did not enhance the expression of HABP2 and HAS2. Kaempferol 3-O-glucoside (K3G) used for a reference compound of 10 also did not enhance HABP2 and HAS2 expression. On the other hand, tiliroside, a reference compound of 12, enhanced the expression of both.
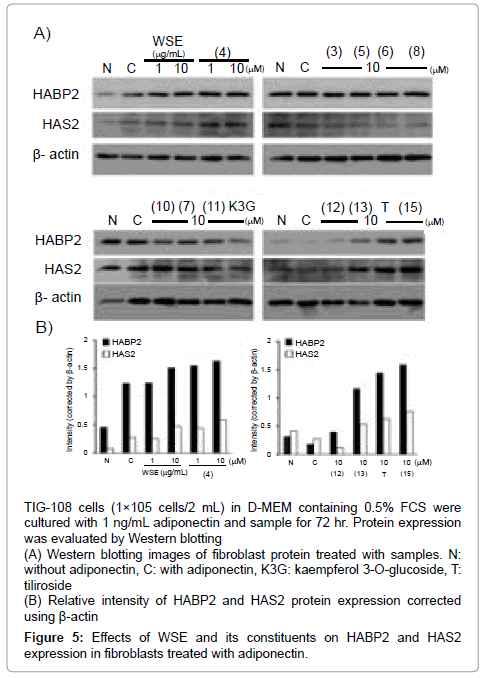
Figure 5: Effects of WSE and its constituents on HABP2 and HAS2 expression in fibroblasts treated with adiponectin.
As a preliminary experiment using subcutaneous adipocytes, we examined the anti-adipogenic effect of test samples in differentiated 3T3-L1 adipocytes. As a result, WSE significantly suppressed lipid accumulation in adipocytes. Among the constituents of water shield, ethyl gallate (3), caffeoyl glucose (5), hypolaetin 7-O-glucoside (6), junsainoside A (12) and kaempferol (15) exhibited suppressive activity at 10 μM. However, specific similarity in their moieties could not be found to exhibit anti-adipogenic activity. Then, the anti-adipogenic effect of WSE and its constituents was evaluated in subcutaneous adipocytes. WSE and its compounds (3, 5, 6, 12 and 15) suppressed lipid accumulation in differentiated subcutaneous adipocytes at a lower concentration than for 3T3-L1 adipocytes. Moreover, ferulic acid (4), kaempferol 3-O-(6"-galloyl) glucoside (7), quercetin 3-O-(6"-galloyl) glucoside (8), quercetin 3-O-glucoside (10) and gossypetin (11) exhibited anti-adipogenic activity at 10 μM. These results suggest that WSE suppresses subcutaneous lipid accumulation effectively compared with that of abdominal adipose tissue and many polyphenols in water shield are involved in the activity.
To evaluate the direct effect of WSE on adipocytokine release, we measured TNF-α and adiponectin released from subcutaneous adipocytes. As WSE enhanced neither TNF-α nor adiponectin production, WSE was found not to affect to adipocytokine release directly. In general, adipocytokine production is changed toward lipid accumulation in adipocytes [22]. Thus, WSE is thought to attenuate adipocytokine release indirectly through the suppression of excessive lipid accumulation in adipocytes. However, high concentrations of 3 and 12 enhanced adiponectin release among selected compounds, showing relatively potent anti-adipogenic activity in subcutaneous adipocytes. Regarding naturally occurring compounds with similar chemical structures to 3 and 12, a green tea catechin with a galloyl moiety (epicatechin gallate) has been reported to increase serum adiponectin in hypertensive rats [23]. On the other hand, two compounds with somewhat similar structures to 12 have been reported to enhance adiponectin production. A citrus polymethoxy flavonoid, nobiletin, enhanced adiponectin secretion from ST-13 adipocytes [24]. Tiliroside, a glycosidic flavonoid with coumaric acid, also enhanced adiponectin production in obese mice [25]. The structure of tiliroside is quite similar to 12. From the activities of these known compounds, 3 and 12 may enhance adiponectin production when applied alone. Galloyl group, methoxyl group in flavonol and cinnamic acid moiety in flavonol glucoside may contribute to the adiponectin release from adipocytes. However, further studies are required to clarify the mechanism of these compounds.
Against the TNF-α-induced fibroblast damage, WSE, ethyl gallate (3) and quercetin (13) suppressed cell damage. TNF-α is released from not only inflammatory cells but also adipocytes. Besides inflamed skin, normal skin TNF-α level is thought to depend on recruitment from capillaries and adipose tissue, and increase with an increment of dermal adipose tissue. Hence, in addition to the anti-adipogenic effect, WSE and its compounds (3 and 13) are thought to be able to control skin TNF-α level.
In several skin disorders, including scleroderma [26], dermatomyositis [27] and psoriasis [28], dermal adiponectin expression has been reported to be lower than in normal skin. Adiponectin expression is also suppressed by morphological changes of the dermis related to abnormal and excessive lipid accumulation, such as cellulite [29] and type 2 diabetes [30]. Thus, adiponectin expression in dermis and subcutaneous adipose tissue is thought to play a crucial role in maintaining normal skin conditions. Upon the examination of hyaluronan binding protein and its synthase, co-treatment of adiponectin with WSE, ferulic acid (4), quercetin (13) and kaempferol (15) enhanced HABP2 and HAS2 expression. As a preliminary examination, we treated fibroblasts with WSE alone. However, WSE did not enhance the expression of HABP2 and HAS2 (data not shown). Thus, WSE and these compounds are thought probably to enhance hyaluronan production synergistically with adiponectin.
In conclusion, WSE and polyphenolic compounds in water shield were found to suppress lipid accumulation in subcutaneous adipocytes. Ethyl gallate and quercetin are involved in the suppressive effect of WSE on TNF-α-induced fibroblast damage. Moreover, ferulic acid, quercetin and kaempferol are found to enhance the expression of HAS2 and HAPB2 with the effect of adiponectin.
This study was not supported by any other foundation except the Oryza Oil & Fat Chemical Co. Ltd.
