Research Article - (2014) Volume 2, Issue 3
Recent trends in anti-aging research projected the use of antioxidant compound derived from botanical products. Phenolic acids, flavonoids and high molecular weight polyphenols are some of antioxidants which are able to protect human skin against the harmful effects of ultraviolet irradiation, i.e. photoaging and skin cancer development. Various studies have demonstrated cocoa beans to contain polyphenols and possess health promoting effects mainly on antiradical property. In this study, the HPLC/DAD quantification of (–)-epicatechin from PBC123 and PBC140 Malaysian unfermented cocoa bean extracts (CBEs) were 121.01 and 118.09 mg/g DW, whereas concentration of (+)-catechin were 6.65 and 6.53 mg/g DW, respectively with no significant differences (p>0.05). In term of antioxidant capacity, Ferric reducing/antioxidant power of the respective clones were assayed at 822.10 and 795.99 mM FeSO4/g DW. Inhibition of proteinases expression induces by reactive oxygen species were exhibited in the anti-collagenase and anti-elastase assays. The anti-collagenase activity of PBC123 and PBC140 were 62.99% and 59.96% whereas anti-elastase has been measured at 36.60% and 15.75%, respectively. Positive and high correlation were observed within epicatechin content (1), FRAP (2) and anti-collagenase (3) with significant relationships for both PBC123 and PBC140 (r12=0.901, r13=0.768 and r23=0.908). A statistical One-Way ANOVA showed that there was no significant difference obtained between PBC123 and PBC140 in terms of epicatechin, FRAP and anti-collagenase assays, however, significant difference was observed from anti-elastase assessment (p<0.05).These results indicate unfermented PBC123 clone as a potential source of natural ingredient in a cosmetic industry.
Malaysia was the fourth largest producer of cocoa worldwide in the early 1990’s [1]. Unfortunately, declining of cocoa bean production in Malaysia since the decade has raised concern of economic viability and sustainability of the cocoa planting industry. Apart from low prices, Cocoa Pod Borer (CPB) infestation and labor issue which were often quoted as reasons for the loss of interest shown in the crop have been identified as the major factors responsible for its decline. Two potential Malaysian cocoa clones, namely PBC140 and PBC123, known for their high yield, i.e. 1,677 and 1,453 kg/ha/year, respectively and high resistance against Vascular Streak Disease (VSD) and CBP infestation, gained lot of attentions for the past few years [2]. Azizah et al. [3] in their study have reported that in accordance to the West African beans, Malaysian beans are traded at a lower price due to inadequate standard of cocoa aroma, astringency and bitter taste characteristics. One of the factors to such occurrence might be due to its high amount of phenolic substances. Moreover, unfermented beans contain significant amount of phenolic content (12-18% w/w) [4] with 60% are comprised of flavanol monomers (epicatechin and catechin) and procyanidin oligomers (dimer to decamer).
Nowadays, the number of natural products that has been used for the development of skin care products is in the increasing trend. Most of these natural products have been proven to exhibit significant antioxidant [5], emollient (María Elena et al.,) [6] as well as for ultraviolet (UV) protection [7] which are crucial for skin care product formulation. The properties are most probably due to the presence of particular phytochemicals, e.g. an antioxidant, in the natural products as active compound. Moreover, natural ingredients are easier to be absorbed by the superficial layers of the skin, thus can lower skin allergy problems. An antioxidant is characterized by any chemical substance which, when observed at relatively low concentration in the body, could decrease or prevent oxidation of substrates [8]. Polyphenols were reported as one of the potent antioxidant besides other common dietary antioxidants, such as ascorbic acid, vitamin E, β-carotene and selenium. Epidemiological studies showed many of the polyphenols exhibit a wide range of biological effects such as an antibacterial, antiviral, anti-inflammatory, anti-allergic, antiradical, anti-aging, vitamin C protection against Reactive Oxygen Species (ROS) and reduction of α-tocopheryl radical. There has been encouraging plant sources associated with polyphenols content like fruits, vegetables [9-11]. Cocoa beans have been said to contain significant amount of polyphenols particularly epicatechin and catechin which were reported as a potential candidate in free radicals scavenging either present in our body system or inner layer of the skin stratum corneum [12].
It is interesting to note that aging can be slowed down, or even stopped for a while and sometimes it can be reversed. Apparently, a study by Lee et al. (2012). on Labisia pumila exhibited that the antioxidant-containing extract worked as a photo protective agent against ROS attacks generated by UV exposure, hence extrinsic aging can be avoided. According to [13], polyphenols have been spectrophotometrically showed high antioxidant properties via free radical scavenging, Ferric Reducing Antioxidant Power (FRAP) and β-carotene bleaching activities and proven to be highly correlated with anti-collagenase and anti-elastase activities. In another study conducted on isolated green tea (Camellia sinensis), polyphenols such as catechin and epigallocatechin gallate have been found to be the inhibitors of collagenase and elastase, subsequently decreasing skin wrinkles [14]. Additionally, cocoa polyphenols have been proven to exhibit a positive action on skin elasticity and skin tonus, namely glycosaminoglycans and collagen I, III and IV [12]. However, the studies of phenolic compounds localizing in Malaysian unfermented cocoa bean extract especially in delaying aging process are limited at presence. Thus, this investigation is aimed on the studies of total phenolics quantification, antioxidant capacity (AOC), anti-collagenase and anti-elastase characteristics of Malaysian unfermented cocoa bean extractas an active ingredient in wrinkle-care cosmetics.
Standards and reagents
Epicatechin, catechin, gallic acid, ascorbic acid, ferrous sulphate (FeSO4•7H2O) and 2,4,6-tri(2-pyridyl)-1,3,5-triazine (TPTZ) were purchased from Sigma Chemical Co. (St. Louis, MO, USA); Folin-Ciocalteu reagent (FC), sodium carbonate, ferric chloride (FeCl3•6H2O) and ethanol were obtained from Merck (Darmstadt, Germany); 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES buffer), N-Isobutyl-N-(4-methoxyphenylsulfonyl)glycyl hydroxamic acid (NNGH), elastatinal, fibroblast collagenase (matrix metalloproteinase-1, MMP-1), Ac-PLG-[2-mercapto-4-methylpentanoyl]- LG-OC2H5, human neutrophil elastase (metalloelastase, MMP-12) and MeOSuc-Ala-Ala-Pro-Val-pNA were purchased from Enzo Life Sciences (Enzo Life Sciences International, Inc., PA, USA).
Plant material
Fresh cocoa beans of Malaysian PBC123 and PBC140 clones were acquired from the Cocoa Research and Development Center, Malaysian Cocoa Board, Jengka, Pahang, Malaysia. The beans were immediately sun-dried for 4 consecutive days without fermentation. The dried and unfermented beans were later manually deshelled prior to grinding.
Ethanolic extraction and High Performance Liquid Chromatography/ Diode Array Detector (HPLC/DAD) determination of epicatechin and catechin
The extract was prepared according to the method of [15], with a slight modification. The seeds were ground in a high-speed laboratory mill until the particle size was reduced to ~1mm. Lipids were removed from 100 g of the ground seeds by extracting three times with 450 mL of hexane. The lipid-free solids were air-dried to yield 45 g of fat-free material. Ground cocoa cotyledons were further extracted with 70% aqueous ethanol for 2 h at 50°C using an orbital shaker (Unimax 1010, Heidolph, Germany). The ratio between samples to extraction medium was 1:25. The extracts were combined, and the organic solvent was removed by rotary evaporator (IKA® RV 10 control, Staufen, Germany) under partial vacuum at 40oC. The aqueous extracts were lyophilized and kept in air-tight container for further studies.
Ten (10) mg of dried sample material was mixed with 5 mL 100% methanol before injected through a reverse-phase C18 SEP-PAK cartridge (Waters Associates, Milford, MA). Epicatechin and catechin retained in the cartridge were eluted with 10 mL 40% aqueous methanol whereas 10 μL of this solution were injected into the HPLC/DAD. The HPLC/DAD included a Waters Associates model 6000-A pump, U6K Universal injector, and Model 450 variable wavelength detector. Separation of epicatechin and catechin were accomplished on a reversephase analytical column with a mobile phase of water:methanol:acetic acid (87:8:5) pumped at a flow rate of 1.5 mL/min. The eluting epicatechin and catechin were detected by monitoring absorbance at 280 nm and quantified by comparing peak area of the sample to those obtained from authentic standards (Sigma, Co. Chemical, St. Louis, USA).
Total phenolic content
The Total Phenolic Content (TPC) was determined using the method developed by [16] with some modifications. The extract (100 μL) was mixed with 0.75 mL of FC reagent (10-fold dilution with distilled water) and allowed to stand at 22°C for 5 min. Then, 0.75 mL of sodium carbonate (60 g/L) solution was added to the mixture. Following 90 min at 22°C, absorbance was measured at 725 nm using a UV-visible spectrophotometer (UV 1601, Shimadzu Co., Ltd., Kyoto, Japan). Gallic acid at concentrations of 0.02-0.1 mg/mL was used as a standard. TPC was expressed as Gallic Acid Equivalent (GAE) per gram of dried sample.
Ferric reducing/antioxidant power (FRAP) assay
The ferric reducing power of the extract was determined based on the reduction of ferric-TPTZ to blue ferrous-TPTZ colored measured at 593 nm. A high FRAP value indicates high antioxidant capacity. First, FRAP reagent was prepared as described by Benzie and Strain (1996). FRAP reagent (1.8 mL) was pipetted into a test tube and incubated at 37°C in a water bath for 10 min. Then, absorbance was taken as t0. Immediately, 100 μL of sample or standard and 100 μL of distilled water were added to the test tube, mixed and incubated at 37°C for 4 min. Then, the absorbance was taken at 593 nm (t4). The FRAP value was calculated based on the equation described by Benzie and Strain (1996) [17]. The reducing potential of the sample extract from unfermented cocoa bean extracts was determined against a standard curve of ferrous sulphate (200-1000 mM) and the FRAP value was expressed as mM FeSO4 equivalents per gram of dried sample.
Collagenase assay
The assay employed was based on spectrophotometric methods reported in the literature [18] with some modifications for use in a 96-well microplate reader. The assay was performed in 50 mM HEPES buffer. MMP-1 was diluted in assay buffer to 2.5% (v/v) whereby required total volume was 20 μL per well. The mixture is warmed to reaction temperature at 37°C shortly before assay. Synthetic chromogenic substrate, (Ac-PLG-[2-mercapto-4-methyl-pentanoyl]-LG-OC2H5) was dissolved in assay buffer to 4% (v/v) and warmed at 37°C as well. The unfermented CBEs of PBC123 and PBC140 were incubated with MMP-1 in buffer for 15 minutes prior to substrate addition to start the reaction. Positive controls were performed with NNGH and ascorbic acid whereas negative controls were performed without any inhibitor. Absorbance at 412 nm was measured immediately after substrate addition and then continuously for 20 minutes in a microplate reader. Experiments were performed in triplicate and percentage of inhibitor activity remains were calculated using the equation below whereas samples ritual is summarized in Table 1.
| Sample | Assay buffer(μL) | MMP-1 [765 mU/μL] (μL) | Inhibitor [6.5 μM] (μL) | Substrate [1 mM] (μL) | Total Volume (μL) |
|---|---|---|---|---|---|
| Blank | 90 | 0 | 0 | 10 | 100 |
| Negative control | 70 | 20 | 0 | 10 | 100 |
| NNGH | 50 | 20 | 20 | 10 | 100 |
| Ascorbic acid | 50 | 20 | 20 | 10 | 100 |
| PBC 123 PBC 140 |
50 50 |
20 20 |
20 20 |
10 10 |
100 100 |
Table 1: Samples ritual for inhibition of collagenase assay.
Inhibitor activity remaining (%) = (Vinhibitor/Vcontrol) x 100
Velocity reaction of inhibitor, Vinhibitor= Absorbanceinhibitor/mininhibitor
Velocity reaction of control, Vcontrol= Absorbancecontrol/mincontrol
Elastase assay
This assay was performed in 100 mM HEPES buffer. MMP-12 and chromogenic substrate, MeOSuc-Ala-Ala-Pro-Val-pNA were dissolved to 1.11% and 10% (v/v) dilution, respectively, in the assay buffer and warmed to reaction temperature (37°C). The unfermented CBEs of PBC123 and PBC140 were incubated with elastase for 10 minutes prior to substrate intervention to start the reaction. The final reaction mixture (100 μL) contained buffer, 22μU/μL elastase, 2mM substrate and 1000 μg/mL of the test extracts. Positive controls were performed with elastatinal and ascorbic acid whereas negative controls were performed without any inhibitor. Absorbance at 405 nm was measured immediately after substrate addition and then continuously for 10 minutes in a microplate reader. The inhibitor activity remaining (%) of elastase is as the same as collagenase determination. Samples ritual is summarized as follows in Table 2.
| Sample | Assay buffer (μL) | MMP-12 [35 mU/μL] (μL) | Inhibitor [6.5 μM] (μL) | Substrate [1 mM] (μL) | Total Volume (μL) |
|---|---|---|---|---|---|
| Blank | 95 | 0 | 0 | 5 | 100 |
| Negative control | 85 | 10 | 0 | 5 | 100 |
| Elastatinal | 65 | 10 | 20 | 5 | 100 |
| Ascorbic acid | 65 | 10 | 20 | 5 | 100 |
| PBC 123 PBC 140 |
65 65 |
10 10 |
20 20 |
5 5 |
100 100 |
Table 2: Samples ritual for inhibition of elastase assay.
Statistical Analysis
Data were expressed as mean ± standard deviation of triplicates. One-way ANOVA (Minitab version 14.0) was used to determine the mean differences for epicatechin and catechin quantification, FRAP, anti-collagenase and anti-elastase activities of PBC123 and PBC140 clones at a significance level of p<0.05. Pearson’s correlation (r-value) was used to determine correlation within epicatechin content, FRAP and anti-collagenase activity.
TPC and profiling
Traditional and hybrid cocoa beans were sampled and phenolic compounds extracted into 70% aqueous ethanol. The total extracted phenolics were then quantified. Table 3 demonstrates the TPC, expressed as gallic acid equivalent (mg GAE/g on a dry weight basis) and concentration of epicatechin and catechin determined in 2 Malaysian unfermented CBEs, namely PBC123 and PBC140. The results were compared with fermented cocoa beans from different countries conducted by [3] under the same condition of 70% ethanol aqueous extraction. Phenolic compounds undergo a complex redox reaction with the phosphotungstic and phosphomolybdic acids present in the FC reagent [19]. However, it should also be noted that some chemical group of ascorbic acid, organic acids, sugars, aromatic amines could also react with FC reagent [20]. In term of the TPC, there is no significant difference between PBC123 (200.85 ± 3.85 mg GAE/g DW) and PBC140 (200.79 ± 3.27 mg GAE/g DW) (p> 0.05). On the contrary, TPC measured from both unfermented cocoa beans of PBC123 and PBC140 significantly exhibited enormous values when compared to the ones from fermented beans, regardless of the countries of origin (Table 3). Both of the unfermented clones showed as high as 200-fold of TPC in comparison to the fermented beans. The high content of TPC in unfermented cocoa beans of PBC123 and PBC140 might account for the better results found for their antioxidant capacity. Several studies have revealed that the phenolic content in plants are associated with their antioxidant capacities, probably due to their redox properties, which allowed them to act as reducing agents, hydrogen donors, and singlet oxygen quenchers [21]. In general, our data indicate that the TPC of unfermented cocoa bean extracts could serve as a significant source for natural antioxidant.
| Sample | Total Phenolic Content (mg GAE/g DW) | Epicatechin Content (mg/g DW) | |
|---|---|---|---|
| *Fermented **Unfermented |
Malaysian Ghanaian Cote d’Ivoire Sulawesian Malaysian PBC123 Malaysian PBC140 |
0.11 ± 0.01b 0.09 ± 0.02b 0.08 ± 0.04b 0.09 ± 0.03b 200.85 ± 3.85a 200.79 ± 3.27a |
8.2 ± 0.12b 3.0 ± 0.16b 2.8 ± 0.05b 12.2 ± 0.07b 121.01 ± 2.19a 118.09 ± 1.14a |
Table 3: Total phenolic content expressed as mg gallic acid equivalent/g dry weight and epicatechin content in fermented and unfermented cocoa beans from different countries.
Whilst TPC may be a useful indicator of potential nutritional benefit, the actual profile of phenolics within the bean is likely to be more important. Thus, profile of the extractable phenolics was determined by HPLC/DAD. Epicatechin and catechin content were identified by its chromatographic behavior and UV spectra (280 nm). The concentration of epicatechin in PBC123 and PBC140 were 121.01 ± 2.19 and 118.09 ± 1.14 mg/g DW, respectively whereas catechin content determined in the respective clones were 6.65 ± 0.88 and 6.53 ± 0.09 mg/g DW. It is well established that epicatechin is the main polyphenols found in cocoa beans [3,22,23]. For comparison, the content of epicatechin in Malaysian unfermented cocoa beans was at least 14-, 39-, 42- and 10-fold higher than the Malaysian, Ghanaian, Cote d’Ivoire and Sulawesian fermented cocoa beans, respectively (Table 3). A One-Way ANOVA for epicatechin showed that, there was no significant difference obtained between PBC140 and PBC123 (p>0.05). Epicatechin is a major component of the polyphenols in cocoa beans and it is a monomer of procyanidins. Approximately, 35% of the TPC in unfermented cocoa beans is constituted of epicatechin. Unfermented refers to the beans that are sun-dried without fermentation or fermented only 1 to 2 days. Fermentation process will lower the polyphenol content through oxidation and exudation, thus, reducing astringency and bitterness [24]. Furthermore, 6–17% of epicatechin is degraded during fermentation process in a study conducted by Nazaruddin et al. (2006) [25]. Therefore, unfermented cocoa beans would rather be chosen in this study due to its high epicatechin content as compared to fermented beans regardless of the quick process being encountered.
| Epicatechin | FRAP | Anti-collagenase | Anti-elastase | |
|---|---|---|---|---|
| Epicatechin FRAP Anti-collagenase Anti-elastase |
1 0.901* 0.768* 0.631* |
1 0.908* 0.383ns |
1 0.064ns |
1 |
Table 4: Pearson’s correlation coefficients (r) among the analyzed parameters.
Antioxidant capacity based on FRAP assay
FRAP assay measures the reducing potential of an antioxidant reacting with a ferric tripyridyltriazine (Fe3+–TPTZ) complex and producing a colored ferrous tripyridyltriazine (Fe2+–TPTZ) [17]. Generally, the reducing properties are associated with the presence of compounds which exert their action by breaking the free radical chain and donating a hydrogen atom. The reduction of Fe3+–TPTZ complex to blue-colored Fe2+–TPTZ occurs at low pH which can be detected at 593 nm. Figure 1 presents the dose-dependent ferric-reducing powers of sample extracts and the standard (ascorbic acid). Reducing power of all sample extracts and ascorbic acid increased with increasing concentration. The FRAP value of PBC123 and PBC140 were assayed at 822.10 and 795.99 mM FeSO4/g DW, respectively with no significant difference (p>0.05). According to Misnawi, et al. [24], Malaysian beans are well-known to have a low cotyledon pH, hence the high antioxidant capacity of unfermented Malaysian cocoa beans of PBC123 and PBC140 could be due to the highly acidic (low pH) nature of the bean cotyledon, which may influence the pH of the assay medium.
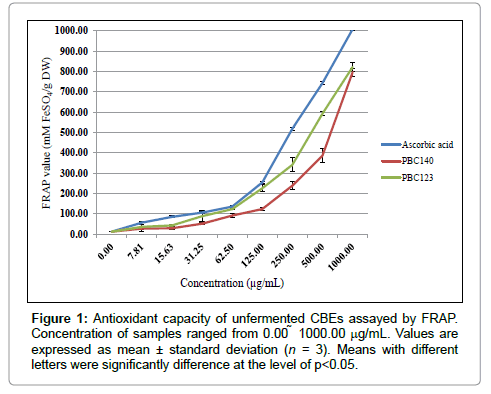
Figure 1: Antioxidant capacity of unfermented CBEs assayed by FRAP. Concentration of samples ranged from 0.00 1000.00 μg/mL. Values are expressed as mean ± standard deviation (n = 3). Means with different letters were significantly difference at the level of p< 0.05.
The reducing power of the studied extracts based on this assay supported the results conducted by Jinap et al. [26]. According to their studies, the unfermented Sulawesian and Malaysian beans which are well-known to have a low cotyledon pH, exhibited the highest antioxidant capacity in comparison to fermented Ghanaian and Cote d’Ivoire beans. Azizah et al. [27] have also found high correlation between the epicatechin and antioxidant capacity of cocoa bean extracts. These findings are in agreement with the present result where the high FRAP value of unfermented PBC123 and PBC140 could have been contributed from the high epicatechin content as shown in Table 3.
Inhibition of collagenase and elastase activities
Collagenase and elastase are known to be major enzymes responsible for dehydration and wrinkle formation on the skin surface. The inhibitory effects of unfermented cocoa bean extracts on these two enzymes are shown in Figure 2. The inhibition of collagenase activities were not significantly different among the NNGH (68.05%), ascorbic acid (64.08%) as well as for PBC123 (62.99%) and PBC140 (59.96%). The elastase inhibitory effects for PBC123 and PBC140 were 36.60% and 15.75%, respectively. This result showed that PBC123 and PBC140 have significantly higher inhibition effect on collagenase rather than on elastase, which could therefore contribute to anti-wrinkle effect. Collagen and elastin are major components of the connective tissue of the skin. Collagen accounts for 70-80% of the skin weight, providing structural stability. However, collagen is known to be rapidly dismantled by collagenase [28]. As collagenase is a zinc-containing metalloproteinase, the epicatechin within the unfermented cocoa bean extract which is also known to be a metal chelator compound, may bind to the Zn2+ ion within the enzyme, thus preventing it from binding with the substrate (collagen embedded within the dermis layer of the skin) [29]. As a result, hydrolyzable collagen can now be avoided and could therefore improve aging skin.
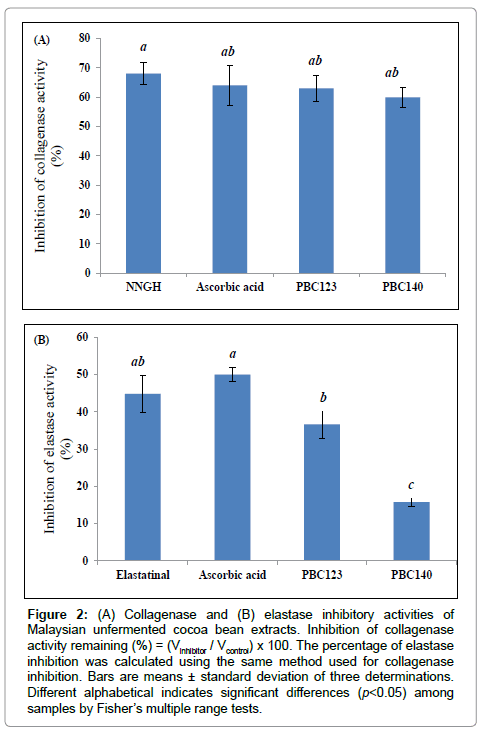
Figure 2: (A) Collagenase and (B) elastase inhibitory activities of Malaysian unfermented cocoa bean extracts. Inhibition of collagenase activity remaining (%) = (Vinhibitor / Vcontrol) x 100. The percentage of elastase inhibition was calculated using the same method used for collagenase inhibition. Bars are means ± standard deviation of three determinations. Different alphabetical indicates significant differences (p< 0.05) among samples by Fisher’s multiple range tests.
Plant phenolic compounds have been known to inhibit collagenase activity [30], accompanied by the reducing power of phenolic compounds to control aging. Cocoa polyphenols, epicatechin and catechin have been proven to exhibit positive action on skin elasticity and skin tonus, namely glycosaminoglycans and collagen 1, III and IV [12]. Moreover, persimmon (Diospyros kaki) leaf-derived polyphenols have showed anti-collagenase and anti-elastase activities [31]. Rosemary (Rosmarinus officinalis) extract has also been known to have strong anti-elastase activity [32]. The principle compound responsible for this inhibitory activity is considered to be the flavonoids present in plant extracts. According to Madhan et al. [33], the conformational change in collagenase induced by the green tea polyphenols is responsible for the inhibition of Clostridium histolyticum collagenase I. In this study, similar kinetic of action was exhibited from the high inhibition of collagenase activity from the Malaysian unfermented cocoa bean extracts of PBC123 and PBC140 (Figure 2A). The hydroxyl part of polyphenols can form hydrogen bond with the functional groups of collagenase. The benzene group of the polyphenols can also be involved in hydrophobic interactions with collagenase.
Elastin, an insoluble fibrous protein, occupies only 2-4% of the dermis matrix, but plays an important role in presenting the elasticity of the skin. It also comprises a network with collagenous fibers under the epidermis [34]. It is usually degraded by elastase, which can hydrolyze peripheral and structural proteins in the connective tissue. Because decomposition of elastin results from activation of elastase caused by the UV light or ROS, inhibition of elastase activity could also be a therapeutic target to protect elastin-induced skin aging.
Our results indicate that PBC123 and PBC140 possess equivalent effect on collagenase, however, the inhibition of elastase by PBC123 (36.60%) was significantly (p<0.05) higher than PBC140 (15.75%). ROS is known to induce expression of proteinases, which cause remodeling of skin extracellular matrix. Activation of such proteases (e.g., matrix metalloproteinases and serine proteases) may be involved in the lack of skin elasticity [35]. The inhibitory effect of PBC123 on elastase could be reflected to our data, which showed slightly higher epicatechin content in PBC123 (Table 3). PBC123 can strongly interact with elastase than PBC140 may be due to hydrophobic interactions, providing the conformational change of elastase, resulting in stronger inhibition of elastase activity. Therefore, our results indicate that PBC123 as a beneficial ingredient in order to treat the effect of aging skin.
Correlations
Finally, correlations were tested to link the different analyzed parameters throughout this study (Table 4). High correlations were found between FRAP values on the one hand and epicatechin (r = 0.901, p< 0.05) or anti-collagenase (r = 0.908, p<0.05) on the other. The FRAP technique used for determining antioxidant capacity has been proven to exhibit high correlation with phenolic compounds in different plants. It has been noted that phenolic compounds, which are known as hydrophilic antioxidants, are secondary metabolites that are mostly abundant in unfermented cocoa bean. Gil et al. [36] found high correlation (r > 0.90, p< 0.05) between antioxidant capacity as determined by FRAP assay and phenolic compounds in nectarines, peaches and plums. Moreover, several studies have found that there were high correlation between total phenolic and antioxidant capacity for cereals [37,38]. A high correlation was also observed between phenolic compounds and antioxidant activity in fruit juices as determined by FRAP or electron spin resonance spectroscopy [39]. In this study, a good correlation between epicatechin and anti-collagenase was found (r = 0.768, p< 0.05) suggested that epicatechin content in unfermented cocoa bean extract has allowed significantly (p< 0.05) increasing level of FRAP, anti-collagenase and anti-elastase activities as exhibited in Table 4. However, inhibition of elastase was not correlated neither with FRAP nor anti-collagenase activity.
In conclusion, the unfermented CBEs of PBC123 and PBC140 are capable of inhibiting collagenase, however elastase is poorly inhibited especially from PBC140. We presume that the enzymes inhibitory effects of unfermented cocoa bean extracts are dependent on their phenolic composition particularly epicatechin content,and molecular structures that define their reactivity towards the ferric reduction potential of sample extracts, as well as on the phenolic concentrations. The epicatechin may bind to the Zn2+ ion within the enzyme, thus preventing it from binding with the substrate and the hydroxyl part of polyphenols which form hydrogen bond with the functional groups of collagenase could therefore inhibit the acceleration of aging skin. Finally, the present study suggests that phenolic compounds of unfermented CBEs provide a good source of natural antioxidant that offer potential protective effect against fibroblast collagen deterioration and which could be exploited by the cosmetics industry.
This research was supported financially by the Ministry of Science, Technology and Innovation of Malaysia (MOSTI) (Project No.: 02-03-13-SF0074).The authors would like to extend their thanks to Universiti Putra Malaysia (UPM) and Chemistry and Technology Division of Malaysian Cocoa Board for the laboratory facilities provided.
