Research Article - (2014) Volume 2, Issue 5
Sambucus formosana (Nakai), a wide spread plant found from the Asia-Pacific regions is used in the traditional medicine of Taiwan to treat trauma induced skin bruise. We herein evaluated in vitro angiogenesis effects of ethanol extract of S. formosana, furthermore, the plant extract effect on HUVEC-mediated tube formation and signaling transduction were also studied. S. formosana extract (SFE) exerted a significant stimulation effect on the proliferation of HUVEC cells at 100 μg/mL. Using migration and invasion assay, the activity of this plant extract on HUVEC was also observed in a dose-dependent manner and significant improved tube formation. Crude extract of S. formosana induced MEK/ERK phosphorylation in HUVEC cells after 10 min of incubation time. Growth factors disturbed assays showed that either VEGF or SFE-induced down stream signaling was inhibited by kinase inhibitors U0126 and PD 98059 and the HUVEC proliferation effect was blocked significantly. These results show that the angiogenesis properties of S. formosana extract seem associated with proliferation of HUVEC cells through down stream regulation of MEK/ERK’s phosphorylation. The regulatory effects of SFE on MEK-ERK and consequently on migration and invasion, and tube formation provide new evidence of its efficacy and emphasize its high potential therapeutic value.
Keywords: Sambucus formosana; HUVEC; Proliferation; Tube formation; MEK/ERK Phosphorylation
Angiogenesis plays an important role in a wide range of physiological processes such as wound healing. A survey of the literatures indicated that some plants and phytochemicals have been used to treat angiogenesis disease [1-4]. Natural products, such as certain Chinese medicines, contain a variety of angiogenic compounds. It has been demonstrated that Rg1, one of the saponins of Ginseng, leads to angiogenesis [1,2]. The angiogenic activity of total saponins from Panax notoginseng was determined by Hong and coworkers [5].
Research achievements suggest the use of plants and their extracts as potential therapeutic agents with pro-angiogenic activity [2]. Globally accepted herbal drugs are thought to be safe and effective, however, more evidence-based confirmation studies are needed. Among the most frequently studied proangiogenic phytochemicals are ginsenosides from Panax ginseng [1], beta-sitosterol from Aloe vera [2], calycosin from Radix Astragali [4], saponins from Panax notoginseng [5] and extracts from Hippophae rhamnoides L. [2], Radix Astragali [6], Puerariae flos [7], Stewartia koreana [8] and Angelica sinensis [9]. Those previous studies are suggested Chinese medicines as formula for treating angiogenesis disease and wound healing. Radix Astragali, Radix Rehmanniae and Angelica sinensis have been combined used and shown to stimulate angiogenesis in some studies also [9,10]; but the mechanism underlying its angiogenic activity remains unclear. Other phytochemicals such as ursolic acid [11], flavonoids [12], ferulic acis [13] and asiaticoside [14] are suggested with angiogenesis effect to promote wound healing in previous studies. All these plant derived stimulators are known to have proangiogenic activies which will benefit human health.
Sambucus formosana Nakai (Caprifoliaceae), one of the blood circulation invigorating herbs, is commonly used in traditional Chinese medicine as blood circulation activation medicine by externally used [15]. The S. formosana is a synonym of S. chinensis Lindl. [16], a widespread shrub in Taiwan, named in Chinese MAO GU XIAO, was one of useful plants in situ. Moreover, leaves of S. formosana are wrapped in a cloth, or poultice made out of leaves and they are applied externally to treat trauma, infectious wounds, and inflammations by Taiwanese aborigines. Several pharmacological effects have been previously reported for Sambucus species, such as, antifungi [17], wound healing [18], osteoblast differentiation [19] and anti-heptatotoxic effect [20]. The activity of S. formosana stimulates differentiation of osteoblasts has been put into light by previous study. Chiou et al. has shown that the chloroform extracts exhibited in vitro differentiation activity in MC3T3-E1 osteoblasts. Wound healing potential of S. ebulus L. extract displayed wound healing activity by in vivo animal studies [18].
The genus Sambucus is widely distributed in Asia. In Taiwan, the root of S. formosana is often used as folk medicine to cure bone fracture, contusion, and pain induced by rheumatism. Traditionally, an alcoholic drench of S. formosana roots and leaves is used for selfmedication. The previous study illustrated that S. formosana with antiinflammatory, anti-nociceptive effect and anti-hepatotoxic protection also [20,21]. Lin and Tome have isolated beta-sitosterol and other triterpene esters from the fresh leaves of S. formosana Nakai [22]. However, at present, there is no direct experimental evidence of the therapeutic benefit of S. formosana in the wounding healing. Therefore, this study was performed to clarify the effect of S. formosana on the proliferation of HUVEC cells. Then the impact of S. formosana on signaling pathways known to be implicated in HUVEC was further explored.
Reagents
Cell culture medium 199 (M199), fetal bovine serum (FBS), and penicillin-streptomycin were purchased from Gibco Life Technologies (Grand Island, NY). Endothelial cell growth supplement (ECGS) was obtained from Upstate (Temecula, CA); Vascular endothelial growth factor (VEGF) was obtained from R&D systems (Minneapolis, MN); Chemicals used here, like heparin, gelatin, Hoechst 33258, sodium dodecyl sulfate (SDS), dimethyl sulfoxide (DMSO), 3-(4,5-cimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), U0126 and PD98059 were purchased from Sigma (St. Louis, MO); Growth factor reduced (GFR) Matrigel™ basement membrane matrix and the BioCoat™ Matrigel™GFR invasion chamber were obtained from BD Biosciences (Bedford, MA). Collagen I Coated Oris™ Cell Migration Assay Kit was obtained from Platypus Technologies (Fitchburg, WI). Cell lysis buffer, antibodies against phospho-VEGF Receptor 2 (Tyr591), VEGF Receptor 2, phospho-Stat3 (Ser727), phospho-Akt (Ser473), phospho-p44/42 MAPK (Erk1/2) (Thr202/ Tyr204), p44/42 MAPK (Erk1/2), phospho-MEK1/2 (Ser217/221), and MEK1/2 (Ser217/221) were purchased from Cell Signaling Technology (Beverly, MA); Immobilon Western Chemiluminescent HRP Substrate was obtained from Millipore (Bedford, MA). Pre-stained protein markers were purchased from Fermentas Life Sciences (Glen Burnie, MD, USA). SuperSignal® West Pico HRP chemiluminescence substrate, the Micro BC™ Protein Assay Reagent protein assay kit and Western Blot Stripping Buffer were from Pierce (Rockford, IL, USA). Nitrocellulose membranes (Hybond-C Extra) were from Amersham Biosciences (Sunnyvale, CA, USA).
Preparation of Sambucus formosana extracts (SFE):
Sambucus formosana Nakai (Caprifoliaceae) was purchased from a Chinese herbal drug store in Taiwan and voucher specimen (0158- 01) of this herbal was deposited in BDL Herbal bank, Taiwan, R. O. C. For extracts preparation, the dried roots and stems of S. formosana (200 g) was pulverized and extracted with 95% ethanol (1L) for 7 days. The combined extracts were evaporated in vacuum to 30 mL and lyophilized. For the cell culture experiments, SFE extracts were dissolved in dimethyl sulfoxide (DMSO) at 200 mg/mL as a stock solution.
HUVEC cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from Cell Applications Inc. (San Diego, CA), and maintained in M199 supplemented with 10% FBS, 100 μ/ml penicillin-streptomycin, 25 μ/ ml heparin, and ECGS on gelatin coated cell culture flasks at 37°C in humidified 5% CO2 incubator. All endothelial cells used in this study were between passages 3 to 6.
HUVEC cell proliferation assay
The cell proliferation effects of SFE on HUVECs were determined by MTT assay and cell counting methods. Cells were seeded at a density of 5×103 cells per well on gelatin-coated 96-well culture plates and incubated in culture medium and allowed to attach for 24 h. The cells were washed twice with M199 and cultured for additional 6 h with M199 containing 2% FBS. Then, the cells were stimulated with various concentrations (10, 30 and 100 μg/ml) of SFE and the control group was treated with 20 mg/mL VEGF or vehicle (0.1% DMSO) for 48 h. Cell proliferation was evaluated by the MTT solution (0.5 mg/ mL) and incubated for another 4 h. 0.1 mL 10% SDS was added to each well to completely dissolve the formazan product and the OD value was detected at the wave length 570 nm. The proliferation ratio was calculated as follows: proliferation ratio (%) = (experimental group OD-vehicle group OD)/(control group OD-vehicle group OD)×100%. In addition, another parallel culture plates were fixed with 10% formaldehyde and the nuclei were stained by using 0.5 μg/mL Hoechst 33258. Fluorescent images were acquired on the fluorescent inverted microscope and the cell number was determined by counting nuclei with Image J software (NIH, USA).
HUVEC cells migration assay
The HUVECs were seeded at a density of 2×104 cells per well onto The Oris™ Cell Migration Assay Collagen I plate and cultured overnight to allow cell adherence to bottom of well. The cells were washed with M199 and cultured for 6 h with M199 containing 2% FBS. Remove Oris™ Stoppers and wash wells to remove unattached cells. Then, the cells were treated with various concentrations (10, 30, 100 and 300 μg/mL) of SFE and the control group was treated with 20 mg/mL VEGF or vehicle. After 18 h incubation, the cells were stained fluorescently with Calcein AM and photographed on each well using EVOS® Microscopes (AMG, Bothell, WA). After Imaging, the cells were fixed in 10% formaldehyde and stained with 0.5 μg/mL of Hoechst 33258. The migrated cell number was determined by counting the number of nuclei with Image J software.
HUVEC cells invasion assay
The HUVECs were seeded at a concentration of 5×104 cells in 500 μL of low serum medium containing various concentrations (10, 30 and 100 μg/mL) of SFE or control group (20 mg/ml VEGF or vehicle) on BioCoat™ Matrigel™ GFR invasion chamber following the manufacturer's instructions. After 18 h incubation, noninvasive cells on the upper surface of the inserts were removed using cotton swabs. The cells were washed and stained with Calcein AM. Fluorescent images of the invasive cells were captured by EVOS® Microscopes at 40x magnification. Then, the inserts were fixed in 10% formaldehyde and stained with 0.5 μg/mL of Hoechst 33258. Invasive cells were quantified by counting the number of nuclei per insert with Image J software also.
Tube formation assay
The HUVECs were harvested and diluted to a concentration of 6×104 cells in 300 μL of low serum medium (M199 containing 2% FBS) with various concentrations (10, 30 and 100 μg/mL) of SFE or control group (20 mg/mL VEGF or vehicle) and seeded on 1:1 M199-diluted GFR Matrigel coated 48-well plate for 3 h. The tube-like structures were photographed by EVOS® Microscopes at 40x magnification. The tube length was measured in three randomly selected fields per well and quantified by Image J software.
Cell lysate preparation and western blot analysis
The HUVECs were seeded in 6-well plates at 2×105 cells per well and starvation in M199 containing 0.1% FBS for 18 h. Then, the cells were stimulated with various concentrations 100 μg/mL of SFE or 20 mg/mL of VEGF. After 10, 30 minutes, 1, 6 or 24 hours, the cells were lysed in cell lysis buffer and scraped off the plates. Cell lysates were quantified and separated by SDS-PAGE (0.1% SDS, 7.5% polyacrylamide gel). After electrophoresis, proteins were transferred to Hyond-C membranes. The membranes were saturated with 5% milk in TBST (50 mM Tris-HCl, 150 mM NaCl, pH 7.5 with 0.1% Tween-20) and probed with primary antibodies against target proteins overnight at 4°C. The membranes were then extensively washed in TBST and probed with corresponding horseradish peroxidase-conjugated secondary antibodies for 1 h. After washing, the signal intensities of target proteins were finally visualized by the Immobilon Western Chemiluminescent HRP Substrate on x-ray film.
Blocking the signaling pathway
The HUVECs were seeded at 5×103 cells/well in a gelatin coated 96-well plate and serum-starved for 24 h. The cells were treated with DMSO (vehicle), 0.5 μM U0126, 5 μM PD98059, 20 mg/mL VEGF or 100 μg/mL SFE. After 48 h, the level of cell proliferation was evaluated. Data are expressed as the percentage cell viability ± SEM of triplicate experiments. For Western blot analysis, the HUVECs were seeded in 6-well plates at 2×105 cells per well and starvation in M199 containing 0.1% FBS for 18 h. Following the pretreatment with kinase inhibitors or not, the cells were stimulated with 20 mg/mL VEGF or 100 μg/mL SFE for another 15 min. These cell lysates were harvested and analyzed by western blot.
Statistical analysis
Data were expressed as means ± the standard deviation (STDEV). The unpaired, two-tailed Student's t test was used to compare the means and STDEV of two independent samples. P-value <0.05 were considered significant. All statistical calculations were performed using Excel 2010 (Microsoft Corporation, Seattle, WA, USA).
SFE stimulates cell proliferation on HUVEC
The effect of SFE on HUVEC proliferation was evaluated using MTT and cell counting assay. Following a 24 h starvation, HUVECs were cultured in low serum medium supplemented with S. formosana extract (10 to 100 μg/mL; 48 h). Cell viability was estimated by determining the amount of formazon product formed in the cell culture medium. As shown in Figure 1a, S. formosana extract promoted cell proliferation in a dose-dependent manner. The maximum increase of cell viability induced by SFE was increased significantly at 100 μg/ ml (P<0.001), compared to vehicle control. A significant (P<0.001) increase in cell proliferation was also observed in VEGF-treated cells, which served as the positive control. But it was not observed in the aqueous fraction of S. formosana extract, SFE-w group). As shown in Figure 1b, S. formosana extract promoted cell proliferation in a dosedependent manner by counting method. To verify the MTT assay results, the cells induced by SFE for 48 h were fixed and stained with H33258. Fluorescent images were acquired on the fluorescent inverted microscope and the cell number was determined by counting nuclei with Image J software. The maximum increase of cell number induced by SFE was proved correspond to MTT assay.
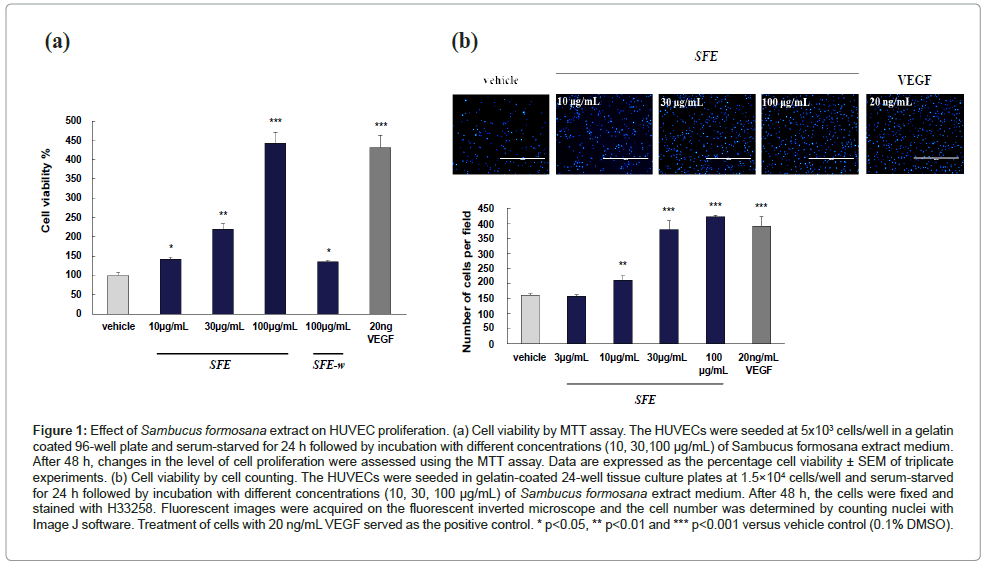
Figure 1: Effect of Sambucus formosana extract on HUVEC proliferation. (a) Cell viability by MTT assay. The HUVECs were seeded at 5x103 cells/well in a gelatin coated 96-well plate and serum-starved for 24 h followed by incubation with different concentrations (10, 30,100 μg/mL) of Sambucus formosana extract medium. After 48 h, changes in the level of cell proliferation were assessed using the MTT assay. Data are expressed as the percentage cell viability ± SEM of triplicate experiments. (b) Cell viability by cell counting. The HUVECs were seeded in gelatin-coated 24-well tissue culture plates at 1.5×104 cells/well and serum-starved for 24 h followed by incubation with different concentrations (10, 30, 100 μg/mL) of Sambucus formosana extract medium. After 48 h, the cells were fixed and stained with H33258. Fluorescent images were acquired on the fluorescent inverted microscope and the cell number was determined by counting nuclei with Image J software. Treatment of cells with 20 ng/mL VEGF served as the positive control. * p<0.05, ** p<0.01 and *** p<0.001 versus vehicle control (0.1% DMSO).
SFE promotes HUVEC cells migration and invasion
The migration of HUVEC was examined using the Oris™ Cell Migration Assay method. The cells migration of the detection zone was measured and compared with the value obtained from vehicle and VEGF treatments. An increase of the migrated cells of detection zone indicated the migration of cells. As shown in Figures 2a and 2b, SFE extract promoted cell migration in a dose-dependent manner. A significant (P<0.001) increase in cell migration was observed in SFE and VEGF-treated group. In the same time, the SFE extract on HUVEC cells invasion was observed and measured by using BioCoat™ GFR Matrigel Invasion Chambers. Quantitative evaluation of the SFE extract induced HUVEC invasion was observed significantly (P<0.001, shown in Figures 2a and 2c). For more focused efforts on wound healing application, the effect of SFE on HUVEC differentiation and vascular formation was assessed by tube formation on Matrigel.
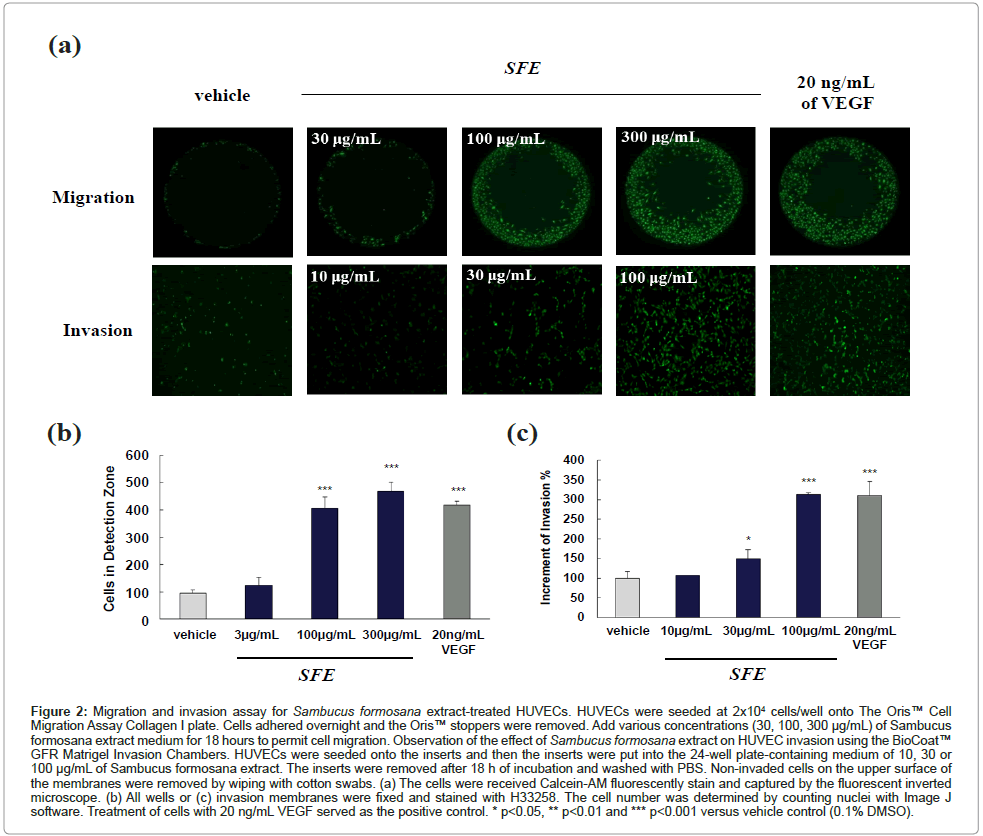
Figure 2:Migration and invasion assay for Sambucus formosana extract-treated HUVECs. HUVECs were seeded at 2x104 cells/well onto The Oris™ Cell Migration Assay Collagen I plate. Cells adhered overnight and the Oris™ stoppers were removed. Add various concentrations (30, 100, 300 μg/mL) of Sambucus formosana extract medium for 18 hours to permit cell migration. Observation of the effect of Sambucus formosana extract on HUVEC invasion using the BioCoat™ GFR Matrigel Invasion Chambers. HUVECs were seeded onto the inserts and then the inserts were put into the 24-well plate-containing medium of 10, 30 or 100 μg/mL of Sambucus formosana extract. The inserts were removed after 18 h of incubation and washed with PBS. Non-invaded cells on the upper surface of the membranes were removed by wiping with cotton swabs. (a) The cells were received Calcein-AM fluorescently stain and captured by the fluorescent inverted microscope. (b) All wells or (c) invasion membranes were fixed and stained with H33258. The cell number was determined by counting nuclei with Image J software. Treatment of cells with 20 ng/mL VEGF served as the positive control. * p<0.05, ** p<0.01 and *** p<0.001 versus vehicle control (0.1% DMSO).
SFE stimulates HUVEC tube formation
The process of angiogenesis is complex, and typically consists of proliferation and alignment to form tubular structures. To test the ability of S. formosana extract to induce HUVEC capillary tube formation, a Matrigel model was used. When HUVECs were cultured on Matrigel – a solid gel of mouse basement membrane proteins – cells aligned easily and formed hollow, tube-like structures. Figure 3 shows that a very low level of tube formation was observed when HUVECs were plated on Matrigel in low-serum medium, whereas morphological changes were observed after treatment with S. formosana extract. Quantitative analysis indicates that S. formosana extract stimulated HUVECs to form more branching points (Figure 3). The numbers of branching points increased in a dose-dependent manner and reached its maximum at a S. formosana extract concentration of 100 μg/mL. A significant (P<0.001) increase in branching points was also observed in VEGF-treated cells, which served as the positive control.
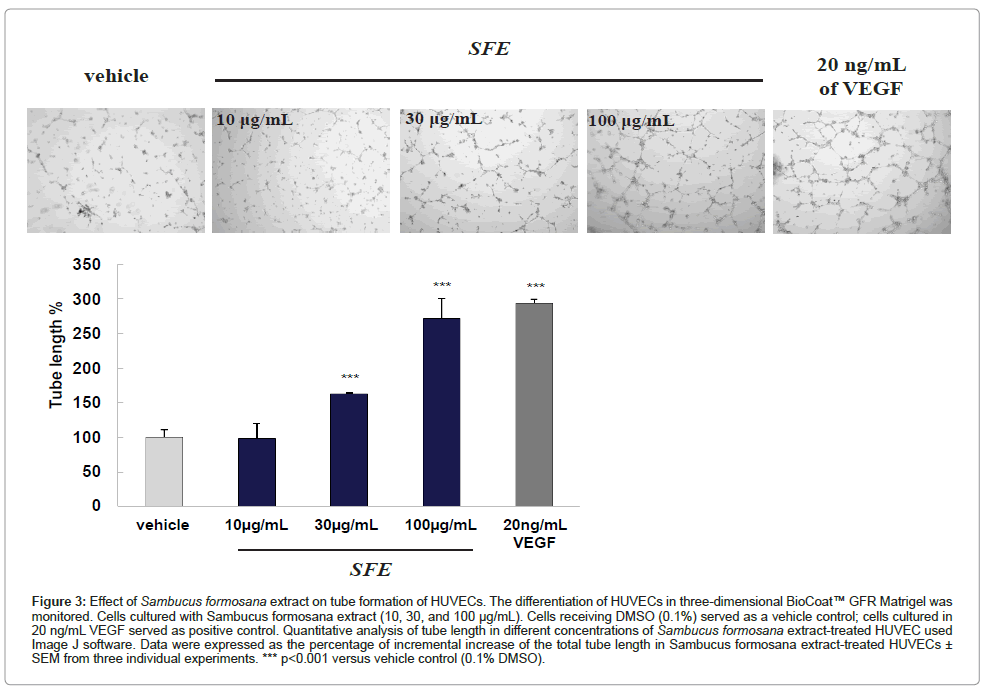
Figure 3: Effect of Sambucus formosana extract on tube formation of HUVECs. The differentiation of HUVECs in three-dimensional BioCoat™ GFR Matrigel was monitored. Cells cultured with Sambucus formosana extract (10, 30, and 100 μg/mL). Cells receiving DMSO (0.1%) served as a vehicle control; cells cultured in 20 ng/mL VEGF served as positive control. Quantitative analysis of tube length in different concentrations of Sambucus formosana extract-treated HUVEC used Image J software. Data were expressed as the percentage of incremental increase of the total tube length in Sambucus formosana extract-treated HUVECs ± SEM from three individual experiments. *** p< 0.001 versus vehicle control (0.1% DMSO).
SFE extract induces angiogenesis via activation of MAPK signaling pathway
ERK1/2, one of the major targets of the MAPK signaling pathway, has been implicated in the regulation of angiogenesis for different functions including cell proliferation, migration and survival. To evaluate the rapid activation of these kinases, western blotting was used to examine the phosphorylation of ERK1/2 following S. formosana extract treatment.
Firstly, phospho-ERK1/2 and total-ERK1/2 were detected following treatment with SFE after different time durations. VEGF stimulated the phosphorylation of ERK1/2 in a time-dependent manner (Figure 4a), which reached a plateau at 30 min, and rapidly declined thereafter. But the SFE stimulated the phosphorylation of ERK1/2 and reached a plateau at 30 min, and sustained to 24 h without declined. It’s totally different from VEGF stimulating signaling. In the same time, the total protein levels of ERK1/2 remained unaffected throughout the course of these experiments.
Furthermore, the phosphorylation of ERK1/2 in HUVECs was enhanced in a dose-dependent manner after incubating with different concentrations of SFE, consistent with the results of the MTT assay (data not shown). To further confirm the involvement of ERK1/2 in S. formosana extract -mediated angiogenesis, a specific blocker was applied to examine its effect on SFE-induced proliferation.
Kinase inhibitors block SFE induced downstream signaling and proliferation
We treated the cells with specific inhibitors of MAPK extracellular signaling-regulated kinase (ERK) kinase (MEK), U0126 and PD98059. And the concentration used here was pre tested that won’t damage HUVEC cells (Figure 4b).
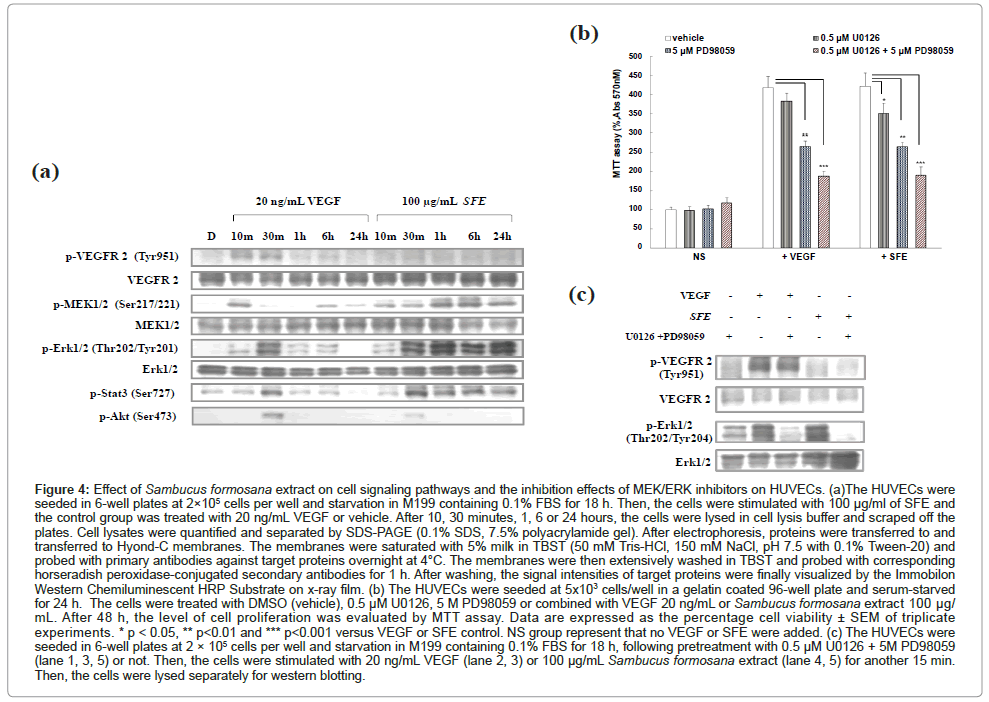
Figure 4: Effect of Sambucus formosana extract on cell signaling pathways and the inhibition effects of MEK/ERK inhibitors on HUVECs. (a)The HUVECs were seeded in 6-well plates at 2×105 cells per well and starvation in M199 containing 0.1% FBS for 18 h. Then, the cells were stimulated with 100 μg/ml of SFE and the control group was treated with 20 ng/mL VEGF or vehicle. After 10, 30 minutes, 1, 6 or 24 hours, the cells were lysed in cell lysis buffer and scraped off the plates. Cell lysates were quantified and separated by SDS-PAGE (0.1% SDS, 7.5% polyacrylamide gel). After electrophoresis, proteins were transferred to and transferred to Hyond-C membranes. The membranes were saturated with 5% milk in TBST (50 mM Tris-HCl, 150 mM NaCl, pH 7.5 with 0.1% Tween-20) and probed with primary antibodies against target proteins overnight at 4°C. The membranes were then extensively washed in TBST and probed with corresponding horseradish peroxidase-conjugated secondary antibodies for 1 h. After washing, the signal intensities of target proteins were finally visualized by the Immobilon Western Chemiluminescent HRP Substrate on x-ray film. (b) The HUVECs were seeded at 5x103 cells/well in a gelatin coated 96-well plate and serum-starved for 24 h. The cells were treated with DMSO (vehicle), 0.5 μM U0126, 5 M PD98059 or combined with VEGF 20 ng/mL or Sambucus formosana extract 100 μg/ mL. After 48 h, the level of cell proliferation was evaluated by MTT assay. Data are expressed as the percentage cell viability ± SEM of triplicate experiments. * p < 0.05, ** p< 0.01 and *** p< 0.001 versus VEGF or SFE control. NS group represent that no VEGF or SFE were added. (c) The HUVECs were seeded in 6-well plates at 2 × 105 cells per well and starvation in M199 containing 0.1% FBS for 18 h, following pretreatment with 0.5 μM U0126 + 5M PD98059 (lane 1, 3, 5) or not. Then, the cells were stimulated with 20 ng/mL VEGF (lane 2, 3) or 100 μg/mL Sambucus formosana extract (lane 4, 5) for another 15 min. Then, the cells were lysed separately for western blotting.
HUVEC proliferation was significantly decreased after pretreatment with ERK activation inhibitor- U0126 and PD98059 (Figure 4b). In the same time, U0126 and PD98059 abrogate VEGF-induced ERK1/2 activation downstream signaling molecules, and disturb HUVEC proliferation significantly (p<0.001). But some proliferation activity remained to support the proliferation effect were observed in VEGF and SFE stimulated group. It was suggested that the phosphor VEGF induced p-STAT3, p-AKT can bypass the MEK/ERK pathway, support the proliferation here. Kinase inhibitor-U0126 and PD98059 block MEK/ERK mediated pathway can’t efficiently stop all the downstream signaling, but the MEK/ERK downstream blocking were determined (Figure 4c). The SFE stimulated signaling was not the same as VEGF induced. In this study, we can’t reveal the upstream involved molecules, but the blocking study shown that U0126 and PD98059 block the MEK/ERK signaling, but not total proliferation function. Some evidence shown here was p-STAT3 in Figure 4a. Altogether, these results indicate that ERK1/2-dependent pathways are involved in SFE-induced HUVEC proliferation, but not the only one involved pathway.
Wound healing occurs as a cellular response to injury and involves activation of fibroblast, endothelial cells and macrophages. Wound healing spans several complicated phases, including inflammation, granulation and re-epithelialization (new tissue formation), contraction and regeneration of tissues [23,24]. Natural products, such as certain Chinese medicines, contain a variety of angiogenic compounds have been reviewed [25-27].
The growth factor modulates diverse processes including proliferation, migration and cell survival. Constitutive mitogenic signaling can be due to overexpression of receptor protein tyrosine kinases or autocrine production of mitogenic growth factors. Ferulic acid was found increase VEGF and platelet-derived growth factor (PDGF) expression in HUVECs [13].
S. formosana Nakai is a plant whose recorded history is indicative of intrinsic wound healing capabilities [15,16]. In the previous study, Lin and Tome isolated beta-sitosterol from the fresh leaves of S. formosana Nakai [27]. The active proangiogenic compound beta-sitosterol has been identified as an angiogenic factor that may be beneficial to the healing process and stimulate migration of HUVEC cells [2]. The wound healing process involves several distinct phases in which the formation of new blood vessels plays an essential role. In this study we describe the angiogenic activity of ethanol extract of the roots and stems of S. formosana Nakai utilizing the HUVEC assay. And also it’s the first investigated SFE induced cell proliferation in HUVEC cells and the activity is comparable to VEGF.
The proliferation activity is related to the ethanol extraction but not aqueous fraction. More subtractions were studied here, but the proliferation effect also diluted (data not shown). The S. formosana extract can be derived from stems or roots, both part of the plants show equivalent activity (data not shown). SFE extracts are demonstrated can promote cell migration and invasion in a dose-dependent manner. More focused efforts on wound healing application, the tube formation effect of SFE on HUVEC differentiation and vascular formation is proved here also. The data demonstrate that S. formosana extract stimulated rapid activation of ERK1/2 in a time- and dose-dependent manner. Our studies provided evidences that S. formosana ethanol extract enhances the proliferation, migration, invasion and tube formation of HUVECs in vitro.
In order to identify the cell signaling mechanism involved in the angiogenic effects of S. formosana ethanol extract, we detected the levels of phosphorylation of MEK1/2, ERK1/2, Stat3, Akt, VEGFR-2 in VEGF or SFE-treated HUVEC signaling transduction. The results showed that 100 μg/mL of SFE significantly activated MEK1/2 and ERK1/2 signaling in 10 minutes, Stat3 and Akt signaling in 30 minutes treatment. However, SFE induced MEK1/2 and ERK1/2 cell signaling transduction can sustain to 24 h, which totally different from VEGF manner.
This study shows that SFE probably stimulates cell signaling pathways to activate the cell proliferation, migration, invasion, and angiogenesis with multiple roles but not via VEGFR-2 phosphorylation. But, whether VEGF or SFE-induced signaling can be blocked by U0126 and PD98059, and disturb HUVEC proliferation significantly. Indicate that MEK1/2 and ERK1/2 pathway play the role in SFE-induced HUVEC signaling.
We report that Sambucus formosana extract (SFE) extracts are demonstrated can promote cell proliferation, migration and invasion in a dose-dependent manner. Our study also provides evidences demonstrate SFE stimulating tube formation, facility HUVEC differentiation and vascular formation, potentially application to wound healing. We suggest that SFE-induced HUVEC proliferation through MEK1/2, ERK1/2 pathway. Blocking this pathway by kinase inhibitor U0126 and PD98059 can disturb HUVEC proliferation significantly. These new evidences of SFE’s efficacy and emphasize its high potential therapeutic value.
This work was supported by the Ministry of Economic Affairs, Taiwan.
