Research Article - (2018) Volume 6, Issue 4
Keywords: Disease severity; Extracts; Fusarium sambucinum; Sampling sites; Sargassum vulgare; Solanum tuberosum L.
Potato (Solanum tuberosum L.) is a strategic crop in Tunisia. Potato-growing areas are estimated at over 23,500 ha ensuring a production of about 385,000 tons [1]. Tubers are stored, from June to November, in refrigerated and or unrefrigerated stores for seed and local market supplies [2].
Fusarium dry rot is one of the main post-harvest diseases of potato worldwide. Serious tuber and seed piece decays may occur during storage leading to heavy losses estimated to up than 25% [3]. Currently, there are at least 20 known Fusarium species responsible for this disease worldwide [4]. Under Tunisian climate conditions, F. sambucinum , F. solani , F. oxysporum f. sp. tuberosi , and F. graminearum were reported as the main species complex involved in the development of Fusarium dry rot with F. sambucinum being the most aggressive one [5-7].
Currently, the primary control for this disease in storage facilities includes the elimination of infected tubers prior to storage and improvement of storage management [8]. Disease control has also been achieved using thiabendazole-based treatments [9]. Unfortunately, repeated use of this active compound together with other benzimidazoles fungicides has led to the development of thiabendazole-resistant strains of F. sambucinum rendering the disease control increasingly difficult [10] especially because no cultivars from all commonly grown potato cultivars in Tunisia were resistant toward Fusarium dry rot [5,6].
Postharvest application of chitosan or β-aminobutyric acid [11], of citric acid or K2HPO4 [12] showed some success against disease development but limited efficacy in commercial storage facilities [13]. Pseudomonas spp. [14], Enterobacter spp. [13], Bacillus spp. [7,15], Trichoderma spp. [16], and non-pathogenic Aspergillus spp. [6] were used for the biocontrol of dry rot incited by F. sambucinum under in vivo or commercial storage conditions. Some plant extracts and essential oils were also explored for the control of the main causal agent of Fusarium dry rot [17,18]. Also, various bioactive compounds were isolated from marine organisms [19].
Sargassum sp., a marine macroalga, is widely distributed on Tunisian coasts [20]. Several researches reported on the broad range of biological activities of Sargassum-derived compounds [21,22]. However, few studies have been published on S. vulgare antifungal activity against phytopathogenic fungi and particularly those infecting potato tubers at the postharvest phase. Therefore, there is an increasing need to develop alternative strategies for controlling major potato storage diseases.
The current study aimed to evaluate the antifungal activity against F. sambucinum of aqueous and organic extracts from S. vulgare sampled on various Tunisian coastal sites.
Fungal inoculum preparation
F. sambucinum isolate was gratefully provided by the Phytopathology laboratory of the Regional Research Centre on Horticulture and Organic Agriculture of Chott-Mariem, Tunisia. It was originally recovered from potato tubers exhibiting Fusarium dry rot symptoms.
Pathogen cultures were initiated from stock cultures maintained at 4°C. They were grown on Potato Dextrose Agar (PDA) amended with streptomycin sulphate (300 mg/L) and incubated at 25°C for one week before being used.
Pathogen virulence was maintained by inoculation of freshly wounded potato tubers, incubation for 21 days at 25°C and re-isolation on PDA medium.
Plant material
Apparently healthy potato tubers cv. Spunta, the most widely grown in Tunisia, were selected for in vivo trials based on their uniformity in size, appearance, ripeness and absence of physical defects. They were previously stored in darkness at 6°C for one month before use. They were brought to room temperature few hours before the beginning of trials. They were thoroughly washed to eliminate adhering soil, superficially sterilized by immersion in a 10% v/v sodium hypochlorite solution for 5 min, rinsed with sterile distilled water (SDW) and air dried.
Algal material sampling and preparation prior to extraction
S. vulgare samples were collected during February 2014 from four different ecological sites, namely Tunis (N 36°51’53.041”; E 10°21’14.4”), Monastir (N 35°46’47.754”; E 10°47’9.312”), Mahdia1 (N 35°30’15.942”; E 11°4’42.035”), and Mahdia2 (N 35°30’13.278” ; E 11°4’34.371”) along the Tunisian coast. They were removed at 1 mdepth beneath the sea surface.
Algal samples were collected by hand, gently washed with seawater and put in plastic bags before being transferred to the laboratory. Collected samples were thoroughly washed several times with tap water, to remove marine epiphytes and sand particles, and shade-dried for three weeks at room temperature. Dry algal materials were grounded into fine powder and stored at 4°C until further use.
Aqueous and organic extraction
Algae collected from the four sampling sites were subjected to aqueous extraction based on Oryan et al. [23] method. Seaweed powder samples of 200 g each were soaked in 2 L of SDW for 24 h under room temperature conditions (25 ± 2°C). Extracts were filtered twice through Whatman N°1 sterile filter paper and further sterilized by filtration through micro-filter (0.22 μm pore size). Collected aqueous extracts were stored at 4°C until further use.
For organic extraction, a 250 g-sample of grounded alga was subjected to a series of maceration in methanol (500 mL) for 2 to 3 days under ambient room conditions as described by Saidana et al. [24]. After filtration, the solvent was evaporated using a rotary evaporator under reduced pressure (at 60°C). The dried algal residue was further subjected to successive extractions using three solvents (350 mL each) with increasing polarities (petroleum ether, chloroform and ethyl acetate). All obtained extracts were filtered and evaporated using a rotary evaporator under reduced pressure and different temperatures depending on the solvent used for the extraction (i.e., 35°C for petroleum ether, 60°C for chloroform and 75°C for ethyl acetate). All dry residues were quantified and individually dissolved into 1 mL of Dimethyl sulfoxide (DMSO) and stored at 4°C until further use.
Test of the in vitro antifungal potential of S. vulgare extracts against F. sambucinum
Aqueous and organic extracts were evaluated for their antifungal activity against F. sambucinum on PDA medium using the poisoned food technique. Appropriate amounts (1-100 mg/mL) of each extract (from the four different sampling sites) were added to molten PDA medium. Extract-amended PDA was aseptically poured into Petri dishes (9 cm in diameter). SDW (aqueous extracts) and DMSO (organic extracts) were used as negative controls. After medium solidification, three agar plugs (6 mm in diameter), cut from 7-day-old cultures, were equidistantly placed in each dish. There were three replicate plates for each individual treatment and the whole experiment was repeated at least twice.
The mean diameter of F. sambucinum colony was measured after 4 days of incubation at 25°C. The percentage of mycelial growth inhibition was calculated as described by Tiru et al. [25].
I=(C-T/C) × 100
where I:Pathogen growth inhibition (in %), C:Colony diameter in control plates, and T:Colony diameter in extract-amended plates.
Test of the fusarium dry rot-suppressive ability of S. vulgare extracts
Potato tubers were wounded at 6 mm depth at two sites along the tuber longitudinal axis using a sterile cork borer. Algal treatments were applied 2 h before pathogen challenge by injecting 100 μL of each extract (aqueous and organic extracts obtained from the four sampling sites and applied at different concentrations) in the occasioned wounds. Tuber inoculation was made by deposing in the occasioned wound a 6 mm-agar plug colonized by F. sambucinum removed from a 7-day-old culture at 25°C.
Negative control tubers were not inoculated with pathogen but similarly treated using only SDW. Positive control tubers were inoculated with F. sambucinum and treated with SDW. Five potato tubers (with two inoculation sites each) were used for each individual treatment. After treatment and pathogen challenge, tubers were incubated for 21 days at 25°C under relatively high humidity.
Fusarium dry rot severity was estimated based on the external and internal extent of the induced decay. Diameters of external rot lesions were recorded and the mean diameter was calculated for each inoculation site. Then, tubers were cut along the longitudinal axis and across the inoculation site to measure rot width (W, mm) and depth (D, mm). Penetration (P, mm) of rotted tissues was estimated using the following Lapwood et al. [26] formula where
P=[(W/2)+(D-6)]/2.
Determination of total phenolic content
Spectrophotometric determination of the total phenolic content in the different organic extracts tested was performed using the Folin- Ciocalteu colorimetric method (according to Benariba et al. [27]).
An aliquot (100 μL) of each S. vulgare organic extract (prepared at two concentrations 1 and 100 mg/mL) was added to 2 mL of a 2% Na2HCO3 solution (w/v). The mixture was shaken and allowed to stand for 5 min before the addition of 100 μL of Folin-Ciocalteu reagent (0.2 N). The solution was mixed thoroughly and incubated for 30 min in the dark and at ambient temperature. After incubation, the absorbance was read versus a prepared blank. The standard curve was prepared using different concentrations of gallic acid.
The total phenol content of the tested extracts was expressed as milligrams of gallic acid equivalents per gram of dry extract (mg gallic acid equivalent/g of extract) from a calibration curve with gallic acid (absorbance vs. concentration of standard). All samples were analyzed in three replicates.
Statistical analyses
Data were analyzed using Statistical Package for the Social Sciences (SPSS) software for Windows version 20.0. They were subjected to oneway analysis of variance (ANOVA) according to completely randomized factorial designs. Means were separated using LSD or Duncan’s Multiple Range tests at P ≤ 0.05.
Pearson’s correlation analysis was performed between total phenolic contents and pathogen growth inhibition potential.
Antifungal potential of S. vulgare aqueous extracts against F. sambucinum
ANOVA analysis showed a highly significant (P ≤ 0.01) inhibitory effect of S. vulgare aqueous extracts against F. sambucinum growth, recorded after 4 days of incubation at 25°C, depending on algal sampling sites, tested concentrations, and their interactions. All aqueous extracts, regardless the sampling sites and concentrations, had significantly suppressed F. sambucinum growth, by 3.56-38.83% as compared to control, excepting those from the site of Tunis applied at 5-25 mg/mL.
Data shown in Figure 1 revealed that all aqueous extracts inhibited F. sambucinum in a concentration dependent manner. Indeed, aqueous extract of S. vulgare sampled from Tunis was found to be more active at 100 and 50 mg/mL where pathogen growth was inhibited by 24.27 and 20.06% respectively, relative to control, compared to 3.56-7.77% recorded with the other concentrations.
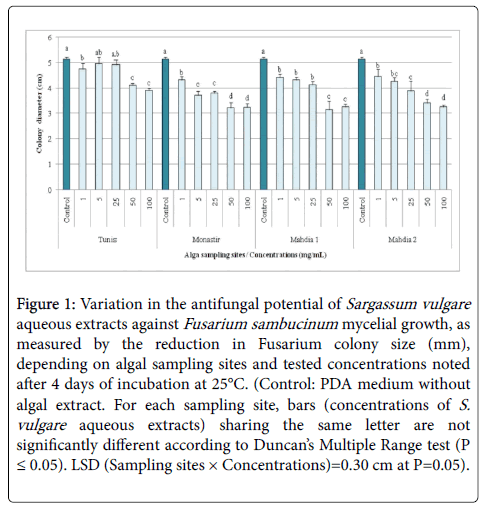
Figure 1: Variation in the antifungal potential of Sargassum vulgare aqueous extracts against Fusarium sambucinum mycelial growth, as measured by the reduction in Fusarium colony size (mm), depending on algal sampling sites and tested concentrations noted after 4 days of incubation at 25°C. (Control: PDA medium without algal extract. For each sampling site, bars (concentrations of S. vulgare aqueous extracts) sharing the same letter are not significantly different according to Duncan’s Multiple Range test (P ≤ 0.05). LSD (Sampling sites × Concentrations)=0.30 cm at P=0.05).
For algae collected from Monastir, the highest inhibition (37%) was noted with extracts applied at 100 and 50 mg/mL compared to 16.18-27.83% achieved at lower concentrations.
Similarly, S. vulgare extract from Mahdia1 was more active at 100 and 50 mg/mL leading to a 36.57-38.83% decrease in pathogen growth compared to 14.24-19.74% noted at 1 and 25 mg/mL. The highest inhibition (33.66-36.57%) was recorded with the highest concentrations used for S. vulgare aqueous extract sampled from Mahdia2 whereas F. sambucinum growth was lowered by 13.59-24.60% with the other concentrations.
It should be highlighted that for all concentrations combined, aqueous extract of S. vulgare sampled at Monastir was the most bioactive against target pathogen, followed by those from Mahdia1, Mahdia2, and Tunis. Moreover, when all sampling sites were combined, S. vulgare aqueous extracts applied at 50 and 100 mg/mL showed comparable efficacy (32.52 and 33.66%) against F. sambucinum growth.
Antifungal potential of S. vulgare organic extracts against F. sambucinum
ANOVA analysis revealed a highly significant (P ≤ 0.01) inhibitory effect of S. vulgare organic extracts against F. sambucinum growth depending on algal sampling sites, the extraction solvent, the tested concentrations and their interactions. All tested organic extracts had significantly reduced F. sambucinum in vitro growth, by 12.97-37.75% compared to control.
For algae sampled on Tunis coasts, pathogen growth was inhibited by 21.61-36.02%, depending on concentrations, with petroleum ether extract showing the highest inhibition. However, inhibitions induced by methanol, chloroform and ethyl acetate extracts were estimated at 16.14-31.70, 19.59-21.61, and 17.00-23.34%, respectively (Table 1). Tested at 100 mg/mL, S. vulgare petroleum ether extract had suppressed F. sambucinum growth by 36.02%, relative to control, compared to 21.61% recorded at 1 mg/mL.
| Sampling site |
Tunis | Monastir | Mahdia 1 | Mahdia 2 | ***Average per concentration | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Extract/ Concentration (mg/ml) |
PE | M | C | EA | PE | M | C | EA | PE | M | C | EA | PE | M | C | EA | |
| Control | 5.78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78 a | 5. 78a |
| 1 | 4.53 b (21.61) | 4.85 b (16.14) | 4.55 b (21.32) | 4.80 b (17.00) | 4.43 b (23.34) | 4.82 b (16.71) | 4.63 b (19.88) | 4.88 b (15.55) | 4.37 b (24.50) | 5.03 b (12.97) | 4.62 b (20.17) | 4.97 b (14.12) | 4.45 b (23.05) | 4.57 b (21.04) | 4.15 b (28.24) | 4.92 b (14.99) | 4.66 b (19.42) |
| 5 | 4.33 b (25.07) | 4.60 b (20.46) | 4.55 b (21.32) | 4.55 bc (21.32) | 4.27 bc (26.22) | 4.85 b (16.14) | 4.67 b (19.31) | 4.83 b (16.43) | 4.48 b (22.48) | 4.98 bc (13.83) | 4.37 b (24.50) | 4.93 b (14.70) | 4.43 b (23.34) | 4.72 bc (18.44) | 3.93 b (31.99) | 4.87 b (15.85) | 4.59 c (20.71) |
| 25 | 4.08 c (29.39) | 4.66 b (19.37) | 4.63 b (19.88) | 4.57 bc (21.03) | 4.22 bc (27.09) | 4.65 b (19.60) | 4.72 b (18.44) | 4.78 b (17.29) | 4.42 b (23.63) | 4.85 bc (16.14) | 4.45 b (23.05) | 4.83 b (16.43) | 4.23 bc (26.80) | 4.35 c (24.78) | 3.92 b (32.28) | 4.58 c (20.75) | 4.50 d (22.25) |
| 50 | 3.92 c (32.28) | 4.65 b (19.60) | 4.65 b (19.59) | 4.50 c (22.19) | 4.12 d (28.82) | 4.88 b (15.56) | 4.43 bc (23.34) | 4.48 c (22.48) | 4.32 b (25.36) | 4.82 bc (16.71) | 4.48 b (22.48) | 4.85 b (16.14) | 3.95 cd (31.70) | 4.32 c (25.36) | 3.90 b (32.56) | 4.50 c (22.19) | 4.42 d (23.49) |
| 100 | 3.70 d (36.02) | 3.95 c (31.70) | 4.53 b (21.61) | 4.43 c (23.34) | 3.98 d (31.12) | 4.73 b (18.16) | 4.30 c (25.65) | 4.45 c (23.05) | 3.95 c (31.70) | 4.60 c (20.46) | 4.17 b (27.95) | 4.72 b (18.44) | 3.60 d (37.75) | 3.82 d (34.01) | 3.80 b (34.29) | 4.43 c (23.04) | 4.20 e (27.41) |
| *Average per site and per solvent | 4.39 b | 4.75 a | 4.78 a | 4.77 a | 4.47 c | 4.96 a | 4.76 b | 4.87 ab | 4.55 b | 5.01 a | 4.64 b | 5.01 a | 4.41 c | 4.59 b | 4.25 d | 4.85 a | |
| **Average per site | 4.67 b | 4.76 a | 4.81 a | 4.52 c | |||||||||||||
Table 1: Variation in the antifungal potential of Sargassum vulgare organic extracts against Fusarium sambucinum mycelial growth, as measured by the reduction in the Fusarium colony size (mm), depending on algal sampling sites, extraction solvent and tested concentrations, observed after 4 days of incubation at 25°C.
Pathogen radial growth was decreased by 23.34-31.12% when grown on PDA medium amended with petroleum ether extract of S. vulgare sampled from Monastir followed by chloroform and ethyl acetate extracts (18.44-25.65 and 15.55-23.05%) as compared to 15.56-19.60% noted using the methanol one. Petroleum ether and chloroform extracts of S. vulgare collected on Mahdia1 coast were found to be the most active compared to the two others where the recorded inhibitions ranged between 20.17 and 31.70% when applied at 1-100 mg/mL. However, methanol and ethyl acetate extracts had suppressed F. sambucinum growth by 12.97-20.46 and 14.12-18.44%, respectively, relative to control.
As for samples from Mahdia2, chloroform extract was the most effective against F. sambucinum where the inhibitions rates varied between 28.24 and 34.29% at 1-100 mg/mL. Interestingly, petroleum ether and methanol extracts, tested at 100 mg/mL, had also suppressed F. sambucinum growth by 37.75 and 34.01%, respectively, compared to 23.05-31.70 and 18.44-25.36% achieved with the other concentrations. For ethyl acetate extract, pathogen growth was slowed by 14.99-23.04% depending on tested concentrations.
It should be highlighted that for all types of organic extracts and tested concentrations, those of S. vulgare sampled at Mahdia2 were the most bioactive against F. sambucinum , followed by those from Tunis. Extracts from Monastir and Mahdia1 showed significantly similar growth-suppressive effects. In addition, for the majority of sampling sites and tested concentrations combined, S. vulgare petroleum ether extract showed the lowest F. sambucinum radial growth, followed by the chloroform one.
Furthermore, for all sampling sites and organic extracts combined, antifungal activity varied in a concentration-dependent manner where the highest inhibitions were recorded when extracts were applied at 100 mg/mL (27.41%) compared to 19.42% noted at 1 mg/mL.
Disease-suppressive ability of S. vulgare aqueous extracts
A significant (P ≤ 0.05) variation in Fusarium dry rot severity, as measured by the rot lesion diameter, was recorded, after 21 days of incubation at 25°C, depending on S. vulgare sampling sites, tested concentrations (5-100 mg/mL) and their interactions. All aqueous extracts, whatever sampling sites and concentrations (excepting those from Mahdia1 applied at 5 mg/mL), had significantly reduced rot lesion diameter, by 9.66-47.07% versus the untreated control.
The lesion diameter was reduced by 26.54-47.07% using aqueous extract from Tunis compared to 24.08-32.72, 9.66-35.48, and 30.86-40.31% achieved following tuber treatments with those from Monastir, Mahdia1 and Mahdia2, respectively (Figures 2 and 3).
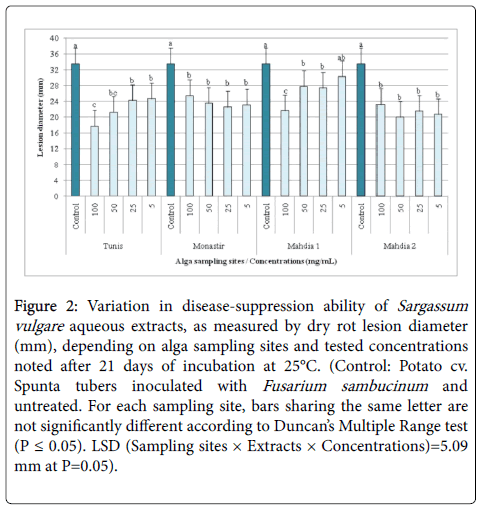
Figure 2: Variation in disease-suppression ability of Sargassum vulgare aqueous extracts, as measured by dry rot lesion diameter (mm), depending on alga sampling sites and tested concentrations noted after 21 days of incubation at 25°C. (Control: Potato cv. Spunta tubers inoculated with Fusarium sambucinum and untreated. For each sampling site, bars sharing the same letter are not significantly different according to Duncan’s Multiple Range test (P ≤ 0.05). LSD (Sampling sites × Extracts × Concentrations)=5.09 mm at P=0.05).
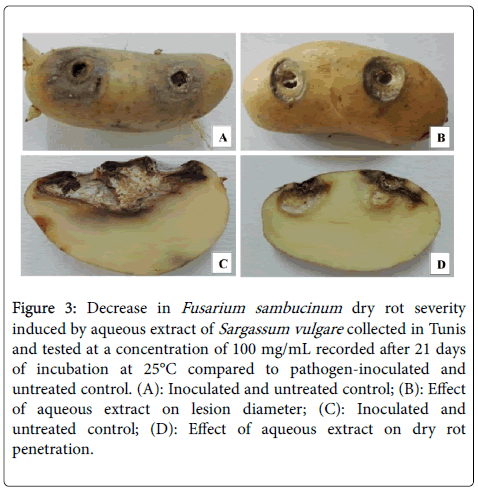
Figure 3: Decrease in Fusarium sambucinum dry rot severity induced by aqueous extract of Sargassum vulgare collected in Tunis and tested at a concentration of 100 mg/mL recorded after 21 days of incubation at 25°C compared to pathogen-inoculated and untreated control. (A): Inoculated and untreated control; (B): Effect of aqueous extract on lesion diameter; (C): Inoculated and untreated control; (D): Effect of aqueous extract on dry rot penetration.
It is worth to note that for all tested concentrations, tubers treated with aqueous extracts of S. vulgare sampled from Mahdia2, Tunis and Monastir sites showed the lowest Fusarium rot lesion diameter (23.78-25.63 mm) as compared to Mahdia1 (28.17 mm). However, when all sampling sites were combined, aqueous S. vulgare extracts exhibited significantly similar efficacy whatever the tested concentrations (varying from 5 to 100 mg/mL) where rot lesion diameter was reduced by 34.37-26.45% relative to control.
In addition, ANOVA analysis performed for the second disease severity parameter, rot penetration, showed a highly significant (P ≤ 0.01) variation depending on used concentrations only as sampling sites had no significant effect on this parameter neither individually nor in interaction with concentrations. In fact, Figure 4 shows that, for all sampling sites combined, aqueous S. vulgare extracts exhibited significantly similar efficacy whatever the concentration tested (varying from 5 to 100 mg/mL) where rot penetration was reduced by 26.54-30.04% versus pathogen-inoculated and untreated control.
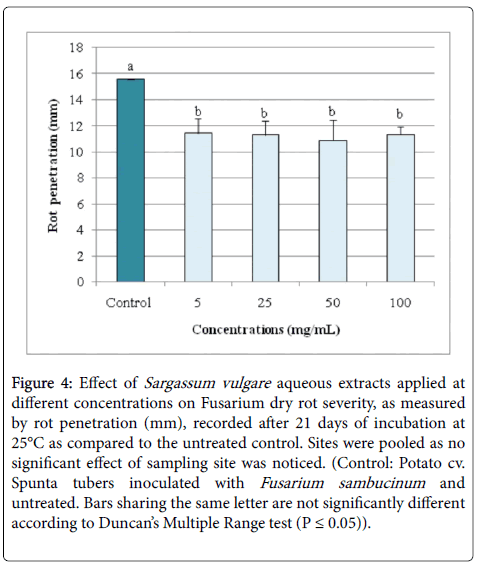
Figure 4: Effect of Sargassum vulgare aqueous extracts applied at different concentrations on Fusarium dry rot severity, as measured by rot penetration (mm), recorded after 21 days of incubation at 25°C as compared to the untreated control. Sites were pooled as no significant effect of sampling site was noticed. (Control: Potato cv. Spunta tubers inoculated with Fusarium sambucinum and untreated. Bars sharing the same letter are not significantly different according to Duncan’s Multiple Range test (P ≤ 0.05)).
Disease-suppressive ability of S. vulgare organic extracts
ANOVA analysis showed that both disease severity parameters, noted after 21 days of incubation at 25°C, varied significantly (P ≤ 0.05) depending on algal sampling sites (Tunis, Monastir, Mahdia1 and Mahdia2), nature of organic extracts (petroleum ether, methanol, chloroform and ethyl acetate extracts), tested concentrations (5-100 mg/mL) and their interactions (two- and three-way interactions). All the tested organic extracts, excepting those from site of Monastir applied at 5-25 mg/mL, those of Mahdia1 tested at 25-100 mg/mL and those of Mahdia2 used at 50-100 mg/mL, were found effective in reducing significantly dry rot severity induced by F. sambucinum , by 5.93-58.29% compared to control, whatever their nature, sampling sites and tested concentrations.
Dry rot lesion diameter was significantly reduced by 42.30-53.18% relative to control, depending on concentrations tested, following treatments with ethyl acetate extract from Tunis compared to 29.55-45.49% achieved with the methanol one. However, treatments based on petroleum ether and chloroform extracts had lowered this parameter by 22.35-36.50 and 17.03-32.60%, respectively (Table 2). For S. vulgare sampled from Monastir, tuber treatment with ethyl acetate extract had reduced lesion diameter by 51.06-54.45% compared to 27.73-40.95, 11.28-18.00, and 9.22-33.72% achieved using methanol, chloroform, and petroleum ether extracts, respectively.
| Sampling site | Tunis | Monastir | Mahdia 1 | Mahdia 2 | ***Average per concentration | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Extract/ Concentration (mg/ml) |
PE | M | C | EA | PE | M | C | EA | PE | M | C | EA | PE | M | C | EA | |
| Control | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a |
| 5 | 31.47 b (25.71) |
25.20 bc (40.52) | 30.36 bc (28.33) | 19.83 c (53.18) | 38.45 ab (9.22) | 28.24 b (33.32) | 34.73 b (18.00) | 19.29 b (54.45) | 34.25 b (19.16) | 29.34 b (30.74) | 27.04 c (36.17) | 24.93 b (41.15) | 36.88 b (12.94) | 31.24 b (26.26) | 28.39 bc (32.97) | 20.97 c (50.49) | 28.79 b (32.04) |
| 25 | 26.90 b (36.50) |
29.84 b (29.55) | 35.15 b (17.03) | 22.80 bc (46.17) | 36.55 ab (13.72) | 30.08 b (28.99) | 37.58 b (11.28) | 20.73 b (51.06) | 39.85 a (5.93) | 26.20 b (38.16) | 31.96 bc (24.55) | 22.55 b (46.75) | 36.21 b (14.52) | 32.65 b (22.92) | 23.37 c (44.82) | 17.67 c (58.29) | 29.38 b (30.64) |
| 50 | 32.53 b (23.20) |
25.92 bc (38.80) | 34.01 bc (19.71) | 21.46 bc (49.34) | 28.07 c (33.72) | 25.01 b (40.95) | 35.23 b (16.84) | 20.11 b (52.52) | 39.02 ab (7.89) | 28.74 b (32.15) | 27.57 c (34.91) | 21.95 b (48.18) | 39.79 ab (6.07) | 27.19 b (35.82) | 35.77 b (15.56) | 25.97 b (38.70) | 29.27 b (30.90) |
| 100 | 32.89 b (22.35) |
23.09 c (45.49) | 28.55 c (32.60) | 24.44 b (42.30) | 32.96 bc (22.19) | 30.61 b (27.73) | 37.27 b (12.02) | 20.16 b (52.41) | 37.40 ab (11.72) | 27.79 b (34.41) | 25.78 c (39.14) | 21.56 b (49.11) | 38.83 ab (8.33) | 29.78 b (29.69) | 33.32 b (21.34) | 26.35 b (37.79) | 29.42 b (30.54) |
| Control | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a | 42.36 a |
| *Average per site and per solvent | 33.23 a | 29.28 b | 34.09 a | 26.18 c | 35.68 a | 31.26 b | 37.43 a | 24.53 c | 38.57a | 30.88 b | 30.94 b | 26.67 c | 38.81 a | 32.64b | 32.64 b | 26.66 c | |
| **Average per site | 30.69 b | 32.23 a | 31.77 ab | 32.69 a | |||||||||||||
Table 2: Variation in the disease-suppression ability of Sargassum vulgare organic extracts, as measured by dry rot lesion diameter (mm), depending on algal sampling sites, extraction solvent and tested concentrations observed after 21 days of incubation at 25°C as compared to the untreated control.
For S. vulgare collected from Mahdia1, ethyl acetate, chloroform and methanol extracts were found to be effective in reducing Fusarium dry rot severity by 41.15-49.11, 24.55-39.14, and 30.74-38.16%, respectively, compared to control whereas only 5.93-19.66% decrease was noted using petroleum ether extract. For samples originating from Mahdia2, ethyl acetate extract had lowered the rot lesion diameter by 37.79-58.29% while 15.56-44.82, 22.92-35.82, whereas a decrease by 6.07-14.52% in this parameter was noted following treatments with chloroform, methanol and petroleum ether extracts, respectively (Figure 5).

Figure 5: Decrease in Fusarium sambucinum dry rot severity (lesion diameter and rot penetration) induced by Sargassum vulgare organic extracts sampled from different Tunisian sites recorded after 21 days of incubation at 25°C compared to pathogen-inoculated and untreated control. (A) (B): Inoculated and untreated control; (C) (D): Ethyl acetate extract tested at 5 mg/mL (Tunis); (E) (F): Ethyl acetate extract tested at 25 mg/mL (Mahdia2); (G) (H): Methanol extract tested at 100 mg/mL (Tunis); (I) (J): Chloroform extract tested at 25 mg/mL (Mahdia2); (K) (L): Petroleum ether extract tested at 50 mg/mL (Monastir).
It should be highlighted that for all organic extracts tested and concentrations used, potato tubers treated with organic extracts of S. vulgare sampled from Tunis and Mahdia1 sites showed the lowest rot lesion diameter as compared to the others. For all sampling sites and concentrations combined, ethyl acetate extract induced the lowest reduction in dry rot severity followed by the methanol one. For all sampling sites combined, organic S. vulgare extracts exhibited significantly similar efficacy whatever their origins and tested concentrations (from 5 to 100 mg/mL) where lesion rot diameter was decreased by 30.54-32.04% compared to pathogen-inoculated and untreated control.
All tested organic extracts were effective in suppressing Fusarium dry rot severity, by 16.55-71.50% versus control, whatever their origin, nature and concentration. Data given in Table 3 showed that treatment of potato tubers with ethyl acetate and methanol extracts of S. vulgare sampled from Tunis led to 56.47-70.66 and 55.41-64.21 decrease in rot penetration, compared to 40.95-54.86 and 32.27-39.77% noted using petroleum ether and chloroform extracts, respectively. Interestingly, treatments with ethyl acetate extracts of algae sampled in Monastir induced a strong suppression of rot penetration, by 61.85-68.89% relative to control, compared to 35.69-41.84, 45.43-53.33, and 22.25-42.41%, recorded on tubers treated with chloroform, methanol and ethyl acetate extracts, respectively. Concerning samples from Mahdia1, methanol and ethyl acetate extracts had suppressed dry rot severity by 53.08-56.94 and 46.84-66.42% compared to, 32.37-39.20 and 28.52-34.59% achieved using chloroform and petroleum ether ones, respectively. Treatments based on ethyl acetate extracts of alga issued from Mahdia2 had suppressed dry rot penetration by 58.10-71.50% compared to 45.76-54.79 and 33.80-55.46% obtained using treatments based on methanol and chloroform extracts, respectively, whereas disease suppression ability displayed by petroleum ether extract was estimated at 16.55-29.21% relative to control (Figure 5).
| Sampling site | Tunis | Monastir | Mahdia 1 | Mahdia 2 | ***Average per concentration | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Extract/ Concentration (mg/ml) |
PE | M | C | EA | PE | M | C | EA | PE | M | C | EA | PE | M | C | EA | |
| Control | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a | 28.82 a |
| 5 | 19.11 b (33.69) | 11.35 b (60.62) | 17.02 b (40.95) | 8.45c (70.66) |
21.56 b (25.19) |
15.26 b (47.05) | 16.76 b (41.84) | 10.99 b (61.85) | 18.85 b (34.59) | 12.64 b (56.12) | 17.52 b (39.20) | 13.93 b (51.65) | 24.05 b (16.55) | 15.42 b (46.48) | 13.95 c (51.57) | 12.07 b (58.10) | 15.56 b (46.01) |
| 25 | 17.90 b (37.89) | 12.85 b (55.41) | 15.11 bc (47.56) | 11.55 b (59.93) |
21.56 b (25.19) |
14.89 b (48.34) | 18.53 b (35.69) | 9.94 b (65.51) | 19.82 b (31.21) | 12.60 b (56.28) | 19.49 b (32.37) | 15.32 b (46.84) | 22.73 b (21.12) | 15.63 b (45.76) | 12.83 c (55.46) | 8.21 c (71.50) | 15.56 b (46.00) |
| 50 | 19.52 b (32.27) | 10.31 b (64.21) | 15.61 bc (45.81) | 12.54 b (56.47) |
16.59 c (42.41) |
13.45 b (53.33) | 18.14 b (37.03) | 8.97 b (68.89) | 19.99 b (30.64) | 12.41 b (56.94) | 19.32 b (32.94) | 9.68 c (66.42) | 20.40 c (29.21) | 13.03 b (54.79) | 16.37 bc (43.18) | 12.00 b (58.37) | 14.90 b (48.31) |
| 100 | 17.36 b (39.77) | 12.18 b (57.72) | 13.01 c (54.86) | 11.66 b (59.55) |
22.20 b (22.95) |
15.72 b (45.43) | 18.21 b (36.81) | 9.20 b (68.07) | 20.60 b (28.52) | 13.52 b (53.08) | 19.06 b (33.86) | 14.34 b (50.22) | 20.64 c (28.36) | 15.19 b (47.30) | 19.08 b (33.80) | 11.78 b (59.10) | 15.86 b (44.96) |
| *Average per site and per solvent | 20.54 a | 15.10 c | 17.91 b | 14.60 c | 22.15 a | 17.63 c | 20.09 b | 13.58 d | 21.61 a | 16.00 b | 20.84 a | 16.42 b | 23.33 a | 17.62 b | 18.21 b | 14.58 c | |
| **Average per site | 17.04 b | 18.36 a | 18.72 a | 18.43 a | |||||||||||||
Table 3: Variation in the disease-suppression ability of Sargassum vulgare organic extracts, as measured by rot penetration (mm), depending on algal sampling sites, extraction solvent and tested concentrations noticed after 21 days of incubation at 25°C as compared to the untreated control.
It should be signaled that for sampling sites and concentrations tested, ethyl acetate and methanol extracts were found to be the most effective for Fusarium dry rot control. Moreover, whatever the concentrations tested, potato tubers treated with extracts of S. vulgare collected from Tunis showed the lowest rot penetration records compared to the other sites.
For all sampling sites combined, organic S. vulgare extracts exhibited significantly similar efficacy whatever the tested concentrations where rot penetration was decreased by 44.96-48.31% as compared to pathogen-inoculated and untreated control.
It should be mentioned that F. sambucinum was successfully reisolated on PDA medium, after 6 days of incubation at 25°C, from all inoculated and treated potato tubers. However, the growth rate of F. sambucinum colonies recovered from tubers previously treated with the different organic extracts was lowered compared to control. Moreover, this recovery and slowed growth were accompanied with a discoloration of pathogen mycelium and a significant variation in its macro-morphological traits. These alterations were more evident following tuber treatments with S. vulgare methanol and ethyl acetate extracts. Furthermore, light microscopic observations revealed an important decrease in pathogen sporulation as compared to control.
Determination of total phenolic content
ANOVA analysis of the tested organic extracts showed a highly significant (P ≤ 0.01) variation of Folin-Ciocalteu total phenolic contents depending on alga sampling sites, types of organic extracts, tested concentrations, and their interactions.
Table 4 showed that for all S. vulgare organic extracts tested, the total phenolic content recorded at 100 mg/mL were higher than those noted at 1 mg/mL. High amounts were recorded for extracts from Tunis with the highest level noted in the chloroform extract with 5.39 mg GAE/g of extract followed by those of petroleum ether and methanol ones by more than 3 mg GAE/g of extract.
| Sampling sites | Tunis | Monastir | Mahdia1 | Mahdia2 | **Average per Extracts and per concentrations | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Concentrations (mg/mL) | 100 | 1 | 100 | 1 | 100 | 1 | 100 | 1 | 100 | 1 |
| Petroleum ether | 3.46 ± 0.10 b | 0.27 ± 0.01 ab | 3.18 ± 0.19 a | 0.22 ± 0.02 b | 3.42 ± 0.17 a | 0.18 ± 0.06 b | 7.56 ± 0.32 a | 0.14 ± 0.02 b | 4.40 ± 0.19 a | 0.20 ± 0.01 a |
| Methanol | 3.01 ± 0.45 b | 0.39 ± 0.01 ab | 3.15 ± 0.15 a | 0.27 ± 0.01 a | 1.36 ± 0.23 bc | 0.18 ± 0.01 b | 2.10 ± 0.01 b | 0.10 ± 0.01 b | 2.40 ± 0.21 b | 0.24 ± 0.02 a |
| Chloroform | 5.39 ± 0.45 a | 0.24 ± 0.02 b | 3.00 ± 0.12 a | 0.13 ± 0.04 d | 1.70 ± 0.10 b | 0.63 ± 0.28 a | 1.16 ± 0.22 b | 0.10 ± 0.03 b | 2.81 ± 0.22 b | 0.28 ± 0.09 a |
| Ethyl acetate | 1.25 ± 0.03 c | 0.41 ± 0.01 a | 1.74 ± 0.02 b | 0.17 ± 0.01 c | 0.68 ± 0.01 a | 0.37 ± 0.01 b | 1.08 ± 0.25 b | 0.34 ± 0.02 a | 1.19 ± 0.08 c | 0.32 ± 0.02 b |
| *Average per sites and per concentrations | 3.28 ± 0.26 a | 0.33 ± 0.03 b | 2.77 ± 0.12 a | 0.20 ± 0.01 b | 0.20 ± 0.01 b | 0.34 ± 0.09 b | 2.97 ± 0.20 a | 0.17 ± 0.01 b | ||
Table 4: Total phenolic content (in mg of Gallic Acid Equivalent (mg GAE) per g extract) determined in the different Sargassum vulgare organic extracts tested.
A statistically equivalent amount (up to 3 mg GAE/g of extract) was determined for petroleum ether, methanol and chloroform extracts from Monastir compared to 1.74 mg GAE/g of extract noted in ethyl acetate fraction.
Petroleum ether extract from Mahdia1 was found to be rich with phenolic compounds with a rate of 3.42 mg GAE/g of extract. The lowest amount was noted with ethyl acetate extract with only 0.68 mg GAE/g of extract.
The highest total phenolic content (up to 7.5 mg GAE/g of extract) was recorded with petroleum ether extract from Mahdia2 compared to 2.10, 1.16 and 1.08 mg GAE/g of extract recorded with methanol, chloroform and ethyl acetate extracts, respectively.
Pearson’s correlation analysis performed for growth inhibition potential and total phenolic contents (for the two tested concentrations) determined in the different organic extracts tested revealed a highly significant and negative correlation (r=-0.541; P=1.2 E-8) between them. For example, petroleum ether extract tested at 100 mg/mL (whatever algal sampling sites) displayed the highest growth inhibition potential against F. sambucinum where phenolic content was relatively high and ranged between 3.18 and 7.56 mg GAE/g of extract depending on algal sampling sites.
Organic extracts had suppressed disease severity (rot lesion diameter and penetration) with a variable degree depending on sampling sites, types of organic extracts and tested concentrations. Total phenolic content (whatever algal sampling sites and extraction solvent) determined at 100 mg/mL (0.68-7.56 mg GAE/g of extract) had impacted with a variable degree the disease suppressive ability of extracts. Pearson’s correlation analysis revealed that lesion diameter severity of organic extracts tested at 100 mg/mL was significantly and positively related (r=0.437; P=0.002) to total phenolic content.
Postharvest management of potato diseases remains a critical challenge. Thus, there is an increased need for tuber protection using effective and environmentally safe alternatives. This study was undertaken to investigate the antifungal potential of the brown alga Sargassum vulgare collected on different Tunisian sites against F. sambucinum growth and dry rot severity.
The present study showed that S. vulgare aqueous extracts had adversely affected, with different levels, the mycelial growth of F. sambucinum . The highest inhibition, by up to 36%, was induced by aqueous extracts from Monastir, Mahdia1 and Mahdia2 applied at 100 and 50 mg/mL. These results are in accordance with our previous findings [28] where Pythium aphanidermatum mycelial growth was also inhibited by up to 28% using 40-50 mg/mL of S. vulgare aqueous extracts. Similarly, aqueous extract of S. wightii , used at 5 mg/mL, is highly active against Rhizoctonia solani [29]. S. myricocystum aqueous extract tested at 10% is active against Colletotrichum falcatum [30]. Inhibition of C. capsici growth reached 33 and 26% using S. ilicifolium and S. wightii water extracts, respectively [31]. However, S. crassifolium and S. oligocystum aqueous extracts had slightly affected the mycelial growth of A. parasiticus , A. niger and Penicillium expansum [32]. Also, Asned and Abbess [33] and Mahianeh et al. [34] showed that aqueous extracts of S. glaucescens , S. swartzii and S. tennirrimum were inactive against F. solani and that of S. wightii , applied at 5 mg/mL, was totally inactive against F. oxysporum , F. solani and Botrytis cinerea [29]. Thus, the biological activity of Sargassum spp. extracts seems to be dependent on algal species, targeted fungal pathogens, collection site and tested concentrations.
In the current study, S. vulgare organic extracts displayed inhibitory activity against F. sambucinum radial growth depending on algal sampling site and tested concentration. Mycelial growth of F. oxysporum , A. ochraceus , Cladosporium cladosporioides and Epicoccum nigrum was also strongly inhibited by cyclohexane extract of S. vulgare applied at 50 mg/mL. However, Khallil et al. [35] found that chloroform, ethanol and ethyl acetate extracts of S. vulgare were slightly active against F. oxysporum whereas, acetone extract was inactive against all the tested pathogens. In addition, mycelial growth of F. monilliforme and A. niger was totally suppressed using 50 mg/mL of S. vulgare methanol and chloroform extracts but these extracts were inactive against A. flavus and P. expansum [36]. Dichloromethanol [37] and methanol [38] extracts of S. vulgare were inactive against P. digitatum and F. solani , respectively. Ethanol extracts of i and S. swartzii , tested at 2 mg/mL, were totally inactive against F. oxysporum , F. solani , M. phaseolina and R. solani [39].
S. vulgare organic extracts showed significant antifungal potential against F. sambucinum mycelial growth with varying levels depending on the type of organic extracts and the concentrations tested. These results are in agreement with several previous studies. In fact, Salem et al. [40] and Mendes et al. [41] found that ethyl acetate is the best solvent for isolation of antimicrobial compounds followed by methanol. In addition, Manilal et al. showed that the highest antifungal activity was obtained with methanol extract [42]. Rattaya et al. demonstrated that methanol and ethanol extracts of S. polycystum were totally inactive against A. niger even when used at 500 mg/mL [43]. Moorthy et al. stated that antifungal activity is pathogen-specific and depends on type of extraction of secondary metabolites, solvents, crude extract concentration and temperature [44]. It could be associated and/or attributed to their differences in chemical nature, polarity, level of solubility of active biomolecules in each used solvent.
Many studies highlighted the antifungal potential of S. vulgare extracts against soilborne plant pathogens and their effectiveness in reducing their mycelial growth under in vitro conditions. In fact, Khallil et al. demonstrated the strong antifungal activity of S. vulgare cyclohexane extract against eight pathogenic fungi including two Fusarium species [35]. Numerous studies showed the presence of a significant variation in the antimicrobial activity when Sargassum spp. extracts were applied at different concentrations [45-47]. Many investigations pointed that the highest in vitro activity was recorded mainly at the lowest concentrations [45,46] and according to our current in vivo investigation, antifungal potential of organic Sargassum extracts was more effective at 5 mg/mL than at 100 mg/mL.
In the present study, aqueous extracts used as tuber treatment for Fusarium dry rot control had significantly limited the rot lesion diameter compared to control. In line with our results, leaf sprays of tomato plants with S. fusiforme extract had strongly decreased the incidence and severity of diseases caused by B. cinerea , Phytophthora infestans and Oidium spp. [48].
S. vulgare organic extracts were also assessed for their ability to control Fusarium dry rot incited by F. sambucinum . Data revealed that the response of potato cv. Spunta to pathogen-inoculation and treatments with S. vulgare extracts varied significantly depending on solvent for extraction, algal sampling sites, and tested concentrations. Ethyl acetate extracts from S. vulgare sampled at Tunis, Monastir and Mahdia2, applied at the lowest concentration tested (5 mg/mL), had lowered dry rot severity by up to 40-50% relative to control. Similarly, Ammar et al. found that methanol extract of S. vulgare , applied at 1 mg/mL, was effective in suppressing by more than 82% Pythium leak caused by P. aphanidermatum [28].
In the present study, methanol and ethyl acetate were found to be the best solvents leading to the extraction of the most effective secondary metabolites with disease- suppression ability. Our findings are in line with those of Lima-Filho et al. [49] and Ammar et al. [50] who showed that organic solvents yield high activities in comparison to water-based extractions. This result could be related to the presence of bioactive metabolites in this macroalgal species which are not soluble in one solvent but might be soluble in another [51]. This variation in the results may be due to difference in species used, time, and place of sample collection. It is clear that the use of organic solvents provides a higher efficiency in extracting active compounds than water-based methods [49,50,52]. González del Val et al. [53] and Lavanya and Veerappan [29] had also selected methanol as solvent for the extraction of antimicrobial compounds from red, green and brown seaweeds.
Phenolic compounds are one of the most widely occurring groups of phytochemicals known by their important biological activities. Determination of the total phenolic contents on S. vulgare organic extracts (at 100 mg/mL) using the Folin-Ciocalteu method showed presence with variable amounts (ranging from 0.68 to 7.56 mg GAE/g of extract) of phenolic compounds depending on algal sampling sites and type of tested extracts. In accordance with our results, Bambang et al. had demonstrated the richness in phenolic compounds for ethanol and hexane extracts recovered from S. filipendula (12.87 and 1.85 mg GAE/g), S. dublicatum (7.87 and 1.82 mg GAE/g), S. crassifolium (6.81 and 1.08 mg GAE/g) and S. binderi (9.09 and 1.14 mg GAE/g) [54].
Several other investigations revealed that total phenolic content was highly affected by the extraction solvent, the method of extraction and especially the genus of the tested alga [43,55].
Numerous studies confirmed that seaweeds can be considered as a source of bioactive compounds with a wide range spectrum of biological activities [47,56]. Various chemical classes, including phenols, indoles, terpenes, acetogenins, fatty acids (satured, monounsatured, polyunsatured) and volatile halogenated hydrocarbons were detected in the ethanol extract of S. vulgare [47,56,57]. Ammar et al. [28] also showed using HPLC-DAD chemical profiling the presence of phenolic acids and flavonoid compounds in the methanol extracts of S. vulgare . These compounds exhibit a wide range of actions on adverse agents present in marine environment [58] that vary depending on intrinsic and extrinsic factors [59]. All of these factors could act on the spatiotemporal regulation on metabolic expression of the active compounds leading to marked qualitative and quantitative variations among similar species at a smaller scale than different species [60].
This study showed for the first time the in vitro and in vivo inhibitory activity of aqueous and organic extracts from S. vulgare collected from Tunisian coastal locations, against F. sambucinum causing potato dry rot. The importance of this work is high as it report on the ability of seaweed extracts to suppress severity of this economically important potato disease and probably other potato pathogens.
