Research Article - (2014) Volume 2, Issue 6
The effect of substrate concentration, pH, temperature and inhibitor (silver nitrate) on the activity of the enzyme invertase was studied using sucrose as the substrate. The invertase acted on the sucrose and converted it to product, glucose and fructose. From Lineweaver-Burk Plot, Vmax was found to be 1 mM/min and the Km approximately 24 mM. At temperature range of 25ËC to 650ËC, the optimum activity was at 450ËC. For a pH range of 3.5-6.0, the pH optimum activity was at 5.5. Inhibition studies showed that silver nitrate inhibited invertase competitively with apparent Km of 150 mM and Ki value of 160 mM.
Keywords: Invertase; Commercial yeast; Confectionery
Enzymes are single-chain or multiple-chain proteins that act as biological catalysts with the ability to promote specific chemical reactions under the mild conditions prevailing in most living organisms [1]. The three distinctive characteristics of enzymes are high specificity, high reaction rate and high capacity for regulation. Enzymes bind reversibly to their substrate at a specific binding site known as the active site. Enzymes accelerate biochemical reaction rates by reducing the energy of activation needed to reach the transition state between reactant and product.
In enzyme catalyzed biochemical reactions, the reactant molecule that binds first to the enzyme is called the substrate [2]. Invertase (β-fructofuranosidase) (E.C.3.2.1.26) is a yeast-derived enzyme and splits sucrose into glucose and fructose.
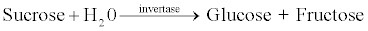
Invertase is mainly used in the food (confectionery) industry where fructose is preferred over sucrose because it is sweeter and does not crystallize easily. Without the aid of invertase, sucrose can be hydrolyzed relatively easily and the reaction proceeds in an acidic environment. There are six classes of enzymes and invertase belongs to the hydrolase class of enzymes. However, the use of invertase is rather limited because, glucose isomerase, can be used to convert glucose to fructose more inexpensively. For health and taste reasons, its use in the food industry requires that invertase be highly purified. Invertase occurs widely in nature and its presence has been reported in plants, certain animal tissues and microorganisms. Invertase or β- fructofuranosidase (EC 3.2.1.26) resulting in the production of invert sugar which has a lower crystal than sucrose at high concentrations, finds numerous applications in the food industry. Confectionary’s preference for invert sugar hovers around its ability to keep the products fresh and soft for prolonged periods. Soluble invertase is used in the sweet industry for the production of artificial honey. Enzyme catalyzed hydrolysis has the advantage of colorless products compared to the colored version obtained through acid hydrolysis [3,4].
A wide range of microorganisms produce invertase and can, thus utilize sucrose as a nutrient. Commercially, invertase is biosynthesized chiefly by yeast strains of Sacharomyces cerevisiae or Sacharomyces carlsbergensis. Even within the same yeast culture, invertase exists in more than one form [5]. For example, the intracellular invertase has a molecular weight of 135,000 Daltons whereas the extracellular variety has a molecular weight of 270,000 Daltons. In contrary to most other enzymes, invertase exhibits relatively high activity over a broad range of pH (3.5-5.5), with the optimum near pH of 4.5. The enzyme activity reaches a maximum at about 55°C. The Michaelis-Menten (Km) values of various enzymes vary widely, but for most enzymes Km is between 2 mM and 5 mM. The Michaelis-Menten value for the free enzyme is approximately 30 mM. Invertase is strongly inhibited by heavy metals. Aniline also inhibits enzyme activity and is considered as a poison. Pesticides or herbicides generally derive their potency from their ability to inhibit enzymes for the growth or survival of organisms [5]. Cures of various diseases are also based on the discovery of enzyme inhibitors.
Substrate and product may also themselves act as enzyme inhibitors. Invertase can be applied for any inversion of sucrose especially liquefied cherry centres, creams, mints, truffles, marshmallow, invert syrup and other fondants. Invertase is used to improve shelf life of confections. It is available in single, double and triple strengths and is packaged in one, ten and 44 pound containers for ease of use, storage and cost efficiency. Invertase 200,000 is powered invertase preparation, which can be applied in the confectionery industry for the production of invert syrup starting from beet or cane sugar. When invertase 200,000 is used for this process, no browning appears and no hydroxymethylfural will form which is a normal phenomenon during hydrolysis with acid. Invertase can be also be used to prevent sugar crystallization in confections by hydrolysis of sucrose (glucose and fructose) in fondants or chocolate coated candies with soft centres. In addition to its main confectionery application, Invertase 200,000 can be used to produce melibiose from raffinose or D-Fructose from inulin, since it contains β - fructosidase activity. It can also be used in some specialty fruit juice products to decrease sucrose level. Invertase appears to be light tan in color, soluble in water and non-flammable. All invertase preparations can be allergens and cause sensitivities on prolonged contact. Gloves and safety glasses should be used when handling this material to prevent skin or eye contact. Repeated exposure can lead to allergic sensitization.
Chandra et al. [6] researched into the complexities of invertase controlling sucrose accumulation and retention in sugar cane. They reported that invertases are involved in a wide variety of metabolic processes affecting plant development. Koch [7] also investigated into the production of thermostable invertases by Aspergillus caespitosus under submerged or solid state fermentation using agro-industrial residues as carbon source. They observed that Aspergillus caespistosus was a new good producer of invertase. Uma et al and Rashad et al [8-10] researched into the production and properties of invertase from Cladosporium cladosporiodies in solid state fermentation (SMF) using pomegranate peel waste as substrate. They concluded that the waste peel could be more efficiently used as a substrate for the production of enzyme under optimized culture conditions.
Effects of substrate concentration on enzyme
When enzyme is mixed with excess of substrate, the initial rate varies hyperbolically with substrate concentration, [S], for a fixed concentration of enzyme. At low substrate concentrations, the occupancy of the active sites on the enzyme molecules is low and reaction rate is related directly to the number of sites occupied. This approximates to first order kinetics in that the rate is directly proportional to the substrate concentration [1]. At high substrate concentrations, effectively all of the active sites are occupied and the reaction becomes independent of the substrate concentration, since no more enzyme-substrate (ES) complex can be formed and zero-order or saturation kinetics are observed. Under these conditions, the reaction rate is dependent upon the conversion of ES complex to products and the diffusion of products from the enzyme. The mathematical equation expressing this hyperbolic relationship between initial rate and substrate concentration is known as the Michaelis-Menten equation.
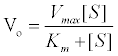
Where Vo=Initial rate
Vmax=Maximum rate of reaction
[S]=Substrate concentration, and
Km=Michaelis constant
Effect of temperature on enzyme
Any environmental factor that disturbs protein structure may lead to a change in enzyme activity. Enzymes are especially sensitive to changes in temperature. All chemical reactions are affected by temperature. In general, the higher the temperature, the higher the reaction’s rate will be. The increase in reaction velocity is due to an increase in the number of molecules that have sufficient energy to enter into the transition rate [11].
The rates of enzyme-catalyzed reactions also increase with increasing temperature. However, enzymes are proteins that become denatured at high temperatures. Each enzyme has an optimum temperature, at which it operates at maximal efficiency. If the temperature is raised to a point somewhat beyond the optimal temperature, the activity of many enzymes decline abruptly. An enzyme optimum temperature is usually close to the normal temperature of the organism it comes from. For example, most human enzymes have temperature optima close to 37°C [11].
Effect of pH on enzyme
Hydrogen ion concentration affects enzymes in several ways. Firstly, catalytic activity is related to the ionic state of the active site. Changes in hydrogen ion concentration can affect the ionization of active site groups. For example, the catalytic activity of a certain enzyme requires the protonated form of a side chain amino group. If the pH becomes sufficiently alkaline that the group loses a proton, the enzyme’s activity may be depressed. In addition, substrates may also be affected. If a substrate contains an ionisable group, a change in pH may alter its capacity to bind to the active site [11].
Secondly, changes in ionisable groups may result in changes in the tertiary structure of the enzyme. Drastic changes in pH often lead to denaturation. Although a few enzymes tolerate large changes in pH, most enzymes are active only within a narrow pH range. For this reason, living organisms employ buffers that closely regulate pH. The pH value at which an enzyme’s activity is minimal is called pH optimum. The pH optima of enzymes vary considerably. For example, the optimum pH of pepsin, a proteolytic enzyme produced in the stomach, is approximately 2. Chymotypsin has an optimum pH of approximately 8.
Effect of inhibitors on enzyme
The activity of enzymes can be inhibited. Studies of the methods by which enzymes are inhibited have practical applications. For example, many clinical therapies and biochemical research tools are based on enzyme inhibition. Enzyme inhibition is an extremely important area of research in the medical field. Lead, mercury, other heavy metals and nerve gases are extremely poisonous to humans because they are inhibitory to enzymes. For example, Pb++ can easily react with the sulphydryl (-SH) groups in a protein. Ag+ ions attach to the histidine side chains of invertase molecule and render it inactive [5].
Inhibition may be reversible or irreversible. Irreversible inhibitors usually bond covalently to the enzyme, often to a side chain group in the active site. In reversible inhibition, the inhibitor can dissociate from the enzyme because it binds through non covalent bonds. Most common forms of reversible inhibition are competitive, non-competitive and uncompetitive inhibition [11]. Some inhibitors of enzymes that form covalent linkages with functional groups on the enzyme are shown below [12] (Table 1).
| Inhibitor | Enzyme groups that combines with inhibitors |
|---|---|
| Cyanide | Fe, Cu, Zn, other transition metals |
| P-Mercuricbenzoate | Sulphhydryl |
| Diisopropylfluoro phosphate | Serine hydroxyl |
| Iodoacetate | Sulphydryl, imidazole, carboxyl, this thioether |
Table 1: Enzyme Inhibitors that form covalent linkages with functional groups.
Materials
Invertase powder from yeast, sucrose, glacial acetic acid, sodium acetate, sodium potassium tartrate, sodium hydroxide and glucose and fructose were obtained from Fluka AG/Chemie, Buchs, Switzerland. Silver nitrate was obtained from Hopkin & Chadwell Health Essex, England and 3,5-dinitrobenzoic Acid (DNB) obtained from BDH Chemical Ltd. Poole, England.
Method
Calibration curve for glucose and fructose: Reaction tubes were set up, as indicated below at 40°C and 1 ml aliquots of 3,5-dinitrobenzoic acid was added into the reaction mixture after 10 minutes to stop the reaction (Table 2). The tubes were placed in a boiling water bath for exactly 5 minutes followed by rapid cooling to room temperature. The absorbance was then read at 540 nm.
| Tube | Absorbance (540 nm) | Concentration (mM) |
|---|---|---|
| 1 | 0.00 | 0.00 |
| 2. | 0.01 | 2.0 |
| 3 | 0.02 | 4.0 |
| 4 | 0.03 | 6.0 |
| 5 | 0.04 | 8.0 |
| 6 | 0.05 | 10.0 |
| 7 | 0.06 | 12.0 |
Table 2: Calibration curve for glucose and fructose.
Effect of enzyme concentration on invertase: Reaction tubes were set up and incubated at 40°C. Aliquots of 1 ml 3, 5-dinitrobenzoic acid (DNB) were added to each reaction mixture after 10 minutes to stop the reaction (Table 3). The tubes were placed in a boiling water bath for exactly 5 minutes followed by rapid cooling to room temperature. The absorbance was then read at 540 nm.
| Tube | Absorbance (540 nm) | Amount of product formed (mM) | Final concentration of invertase (x10-3 mg/ml) |
|---|---|---|---|
| 1 | 0.00 | 0.0 | 0.0 |
| 2. | 0.01 | 2.0 | 0.10 |
| 3 | 0.02 | 4.0 | 0.30 |
| 4 | 0.03 | 6.0 | 0.50 |
| 5 | 0.04 | 8.0 | 0.70 |
| 6 | 0.05 | 10.0 | 0.90 |
| 7 | 0.06 | 12.0 | 1.00 |
Table 3: Effect of enzyme concentration on invertase.
Time course for invertase: Reaction tubes were set up in the time interval of 0-18 minutes as shown below. Aliquots of 1 ml 3, 5-dinitrobenzoic acid (DNB) were added to each reaction mixture after the incubation period to stop the reaction. The tubes were placed in a boiling water bath for exactly 5 minutes followed by rapid cooling to room temperature. The absorbance was then read at 540 nm (Table 4).
| Tube | Time (min) | Absorbance (540 nm) | Product formed (mM/min) |
|---|---|---|---|
| 1 | 0 | 0.000 | 0.0 |
| 2. | 2 | 0.006 | 1.2 |
| 3 | 4 | 0.012 | 2.4 |
| 4 | 6 | 0.018 | 3.6 |
| 5 | 8 | 0.023 | 4.6 |
| 6 | 10 | 0.029 | 5.8 |
| 7 | 12 | 0.035 | 7.0 |
| 8 | 14 | 0.037 | 7.4 |
| 9 | 16 | 0.045 | 9.4 |
| 10 | 18 | 0.057 | 11.4 |
Table 4: Time course for invertase.
Effect of substrate concentration on invertase: The reaction tubes were set up as indicated below and incubated at 40°C. Approximately 1 ml of 3,5-dinitrobenzoic acid were added to each reaction mixture after 10 minutes to stop the reaction. (Table 5). The tubes were placed in a boiling water bath for exactly 5 minutes followed by rapid cooling to room temperature. The absorbance was then read at 540 nm.
| Tube | Absorbance (540 nm) | Product formed (mM) |
|---|---|---|
| 1 | 0.000 | 0.0 |
| 2. | 0.008 | 0.16 |
| 3 | 0.019 | 0.38 |
| 4 | 0.026 | 0.52 |
| 5 | 0.028 | 0.56 |
| 6 | 0.033 | 0.62 |
| 7 | 0.039 | 0.78 |
Table 5: Effect of substrate concentration on invertase.
Effect of pH on invertase: In the determination of optimum pH, reaction tubes were set up in the pH interval of 3.5-6.0 as shown below. Aliquots of 1 ml 3,5-dinitrobenzoic acid (DNB) were added to each reaction mixture after 10 minutes to stop the reaction (Table 6). The tubes were placed in boiling water bath for exactly 5 minutes followed by rapid cooling to room temperature. The absorbance was then read at 540 nm.
| pH | Absorbance (540 nm) | Product formed (mM/min) |
|---|---|---|
| 3.5 | 0.020 | 0.40 |
| 4.0 | 0.023 | 0.46 |
| 4.5 | 0.025 | 0.50 |
| 5.0 | 0.028 | 0.56 |
| 5.5 | 0.033 | 0.66 |
| 6.0 | 0.017 | 0.36 |
Table 6: Effect of pH on invertase.
Effect of temperature on invertase: In the determination of optimum temperature, reaction tubes were set up at temperature interval of 25-65°C as shown below. Aliquots of 1 ml 3,5-dinitrobenzoic acid (DNB) were added to each reaction mixture after 10 minutes to stop the reaction (Table 7). The tubes were placed in boiling water bath for exactly 5 minutes followed by rapid cooling to room temperature. The absorbance was then read at 540 nm.
| Tube | Temperature (°C) | Absorbance (540nm) | Product formed (mM/min) |
|---|---|---|---|
| 1 | 25 | 0.010 | 0.2 |
| 2 | 35 | 0.014 | 0.28 |
| 3 | 45 | 0.03 | 0.60 |
| 4 | 55 | 0.010 | 0.20 |
| 5 | 65 | 0.003 | 0.06 |
Table 7: Effect of pH on invertase.
Effect of an inhibitor (silver nitrate) on invertase: Reaction tubes were set up as below without (a) and with (b) silver nitrate. Aliquots of 1 ml 3,5-dinitrobenzoic acid (DNB) were added to each reaction mixture after 10 minutes to stop the reaction (Tables 8 and 9). The tubes were placed in boiling water bath for exactly 5 minutes followed by rapid cooling to room temperature. The absorbance was then read at 540 nm.
| Tube | Absorbance (540nm) | Product formed (mM/min) | 1/v (mM/min)-1 | 1/[S] (mM/min)-1 |
|---|---|---|---|---|
| 1 | 0.00 | 0.00 | 0.00 | 0.00 |
| 2 | 0.010 | 0.20 | 5.00 | 0.190 |
| 3 | 0.016 | 0.32 | 3.13 | 0.100 |
| 4 | 0.022 | 0.44 | 2.27 | 0.060 |
| 5 | 0.028 | 0.56 | 1.79 | 0.050 |
| 6 | 0.027 | 0.54 | 1.85 | 0.040 |
Table 8: Effect of inhibitor on invertase (without inhibitor).
| Tube | Absorbance (540nm) | Product formed (mM/min) | 1/v (mM/min)-1 |
|---|---|---|---|
| 1 | 0.000 | 0.00 | 0.00 |
| 2 | 0.002 | 0.04 | 25.00 |
| 3 | 0.003 | 0.06 | 16.60 |
| 4 | 0.005 | 0.08 | 10.00 |
| 5 | 0.006 | 0.10 | 8.30 |
| 6 | 0.008 | 0.12 | 6.25 |
Table 9: Effect of inhibitor (with inhibitor, AgNO3).
The results are discussed in the following figures (Figures 1-7).
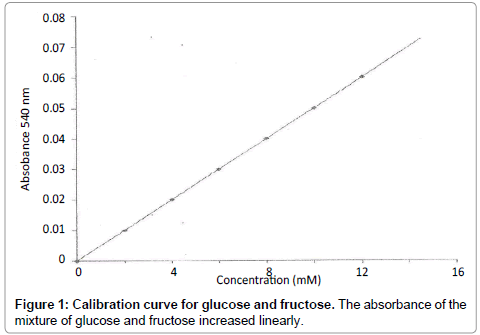
Figure 1: Calibration curve for glucose and fructose. The absorbance of the mixture of glucose and fructose increased linearly.
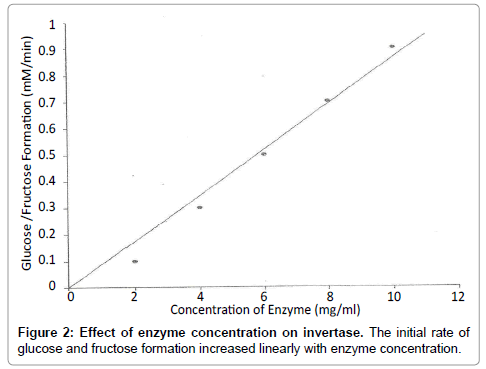
Figure 2: Effect of enzyme concentration on invertase. The initial rate of glucose and fructose formation increased linearly with enzyme concentration.
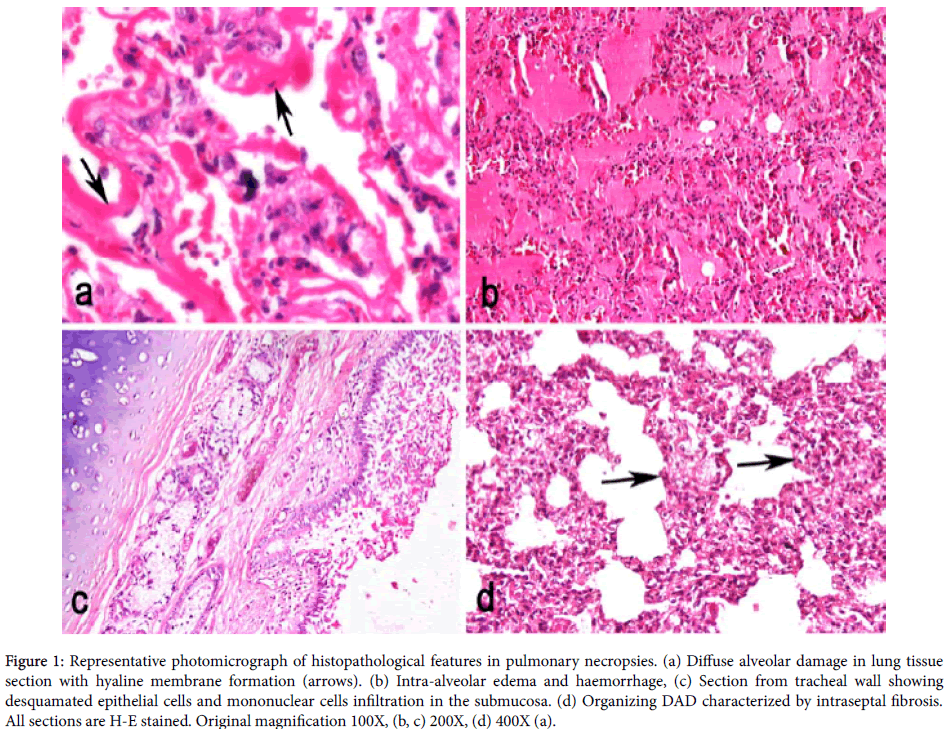
Figure 3: Time course for invertase The formation of glucose and fructose increased linearly with time from 0.0-7.0 mM in a time range of 0-18 minutes.
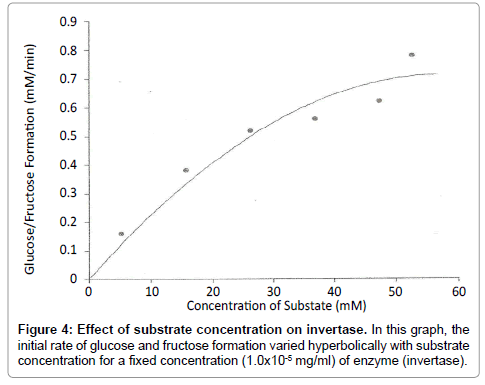
Figure 4: Effect of substrate concentration on invertase. In this graph, the initial rate of glucose and fructose formation varied hyperbolically with substrate concentration for a fixed concentration (1.0x10-5 mg/ml) of enzyme (invertase).
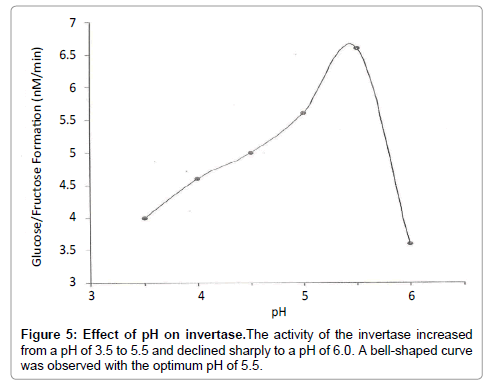
Figure 5: Effect of pH on invertase.The activity of the invertase increased from a pH of 3.5 to 5.5 and declined sharply to a pH of 6.0. A bell-shaped curve was observed with the optimum pH of 5.5.
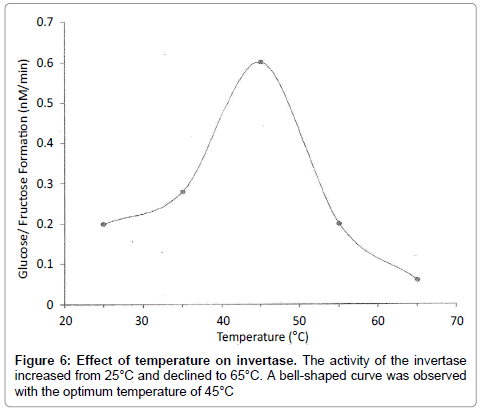
Figure 6: Effect of temperature on invertase. The activity of the invertase increased from 25°C and declined to 65°C. A bell-shaped curve was observed with the optimum temperature of 45°C
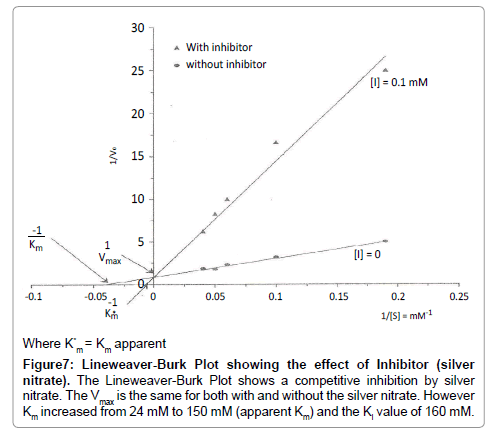
Figure 7: Lineweaver-Burk Plot showing the effect of Inhibitor (silver nitrate). The Lineweaver-Burk Plot shows a competitive inhibition by silver nitrate. The Vmax is the same for both with and without the silver nitrate. However Km increased from 24 mM to 150 mM (apparent Km) and the Ki value of 160 mM.
Discussion
From (Figure 3), the initial rate of reaction for invertase varied hyperbolically with substrate concentration. At low substrate concentrations, approximately first-order kinetics was observed and the initial rate was proportional to the substrate concentration. At high substrate concentrations, zero-order or saturation kinetics existed and the initial rate was independent of substrate concentration. From Line weaver-Burk plot (Figure 7), Km value obtained was 24 mM as compared with that of literature 29 mM [5] and Vmax was 1 mM/min.
In the study of effect of pH on invertase, the pH optimal in the range of 3.5-6.0 was found to be 5.5. The pH of the invertase increased steadily from 3.5 to 5.5. The activity of the invertase declined from pH of 5.5 to 3.5 due to denaturation of the enzyme. The state of ionization of amino acid residues in the active site of the invertase is generally pH dependent. The catalytic activity also relies on a specific state of ionization of these residues. As a consequence, the pH enzyme activity profile is either bell-shaped which represents two important amino acid residues in the active site, giving a narrow pH optimum or a plateau, which represent one important amino acid residue in the active site. In this study, the bell-shape was observed for the pH-enzyme activity profile (Figure 5) and the pH of 5.5 was in conformity with literature review because the enzyme was obtained from the same source.
In the study of effect of temperature on invertase, it was observed that invertase was sensitive to temperature; the higher the temperature, the higher the rate of reaction. The increase in the rate of reaction is due to an increase in the number of molecules that have sufficient energy to enter into the transition state. From (Figure 6), the reaction rate of invertase increased from 25°C to 45°C. The activity of the invertase declined from 45°C to 65°C. This implies that the enzyme was denatured which led to the loss of activity of the invertase. The temperature optimum was therefore found to be 45°C, which is not in conformity with literature value of 55°C. In the study of effect of inhibitors on invertase, it was observed that silver nitrate inhibited invertase comparatively as shown in Figure 7. In competitive inhibition of silver nitrate, it was observed that Vmax (1 mM/min) remains the same whilst Km increased from 24 mM to 150 mM (apparent) and Ki of 160 mM. Probably silver ions bound to the histidine side chain of invertase and rendered it inactive. Ali Jabbar Resheg AL- Sa'ady [13] carried out studies on three Saccharomyces cerevisae isolates from different sources (China, Turkey and Egypt). These isolates were screened by culturing on solid state fermentation to select the most efficient isolate for invertase production. He reported that the isolates from China had high specific activity of 34.7 U/mg and that the optimum pH for enzyme activity was 5.0 whilst that of temperature was reported to be 30°C. Ul-Haq et al. [14] also carried out studies on kinetics of invertase synthesis by Saccharomyces cerevisiae in synthetic medium and reported that the initial concentration of sugar plays an important role in determining the maximum amount of enzyme produced and residual sugars produced after hydrolysis by Saccharomyces cerevisiae [15]. They also observed that the maximum production of invertase was obtained when incubation temperature was maintained at 30°C. Abrahao-Neto et al. [16] also carried out studies on hexokinase production from Saccharomyces cerevisae using studies on the effects of pH (4.0, 4.5 or 5.0), with temperature of (30, 35 or 40°C and dissolved oxygen (DO) (0.2, 2.0, 4.0 or 6.0 mg O2/ L on invertase formation by yeast and reported that setting dissolved oxygen temperature at 4.0 mg O2/L and 35°C respectively, the low reducing sugar concentration accumulated and the high Glucose-6- Phoshate Dehydrogenase (G6PDH )and invertase activities were observed at pH of 4.5. An increase in the pH of 0.5 caused at least a 50% decrease in the activity of invertase and G6PDH.
Conclusion
The Vmax of the conversion of sucrose to glucose and fructose by invertase was 1 mM/min and the Km, was 24 mM. The optimum temperature and pH were found to be 45°C and 5.5 respectively. Inhibition studies showed that silver nitrate inhibited invertase competitively with an apparent Km of 150 mM and a Ki of 160 mM.
