Research Article - (2014) Volume 2, Issue 6
A compilation of the 13C NMR data of the novel aglycones of new triterpenoid saponins, reported during the period mid1996-March, 2007 and arranged skeleton wise is included. The biological activities of the parent saponins are also covered.
Keywords: Triterpenoid saponins; Isolation; Structure elucidation; Biological activity
Triterpenoid saponins are naturally occurring surface active glycosides of triterpenes. They can be divided into two major groups a) monodesmosides in which the aglycone has a singly attached linear or branched chain set of sugars and (b) bisdesmosides in which there are two sets of sugars. They are widely distributed in plant kingdom, certain marine organisms and marine flora and fauna. Their structural diversity, occurrence as complex mixtures, novel bioactivities of relevance to the pharmaceutical industry and agriculture, and usefulness as ingredients of cosmetics, allelochemicals, food and feeding stuff has generated great interest in the study of these molecules. With the introduction of advanced methods of isolation and structure determination there has been increased activity in these molecules. In recent years, the discovery of novel triterpenoid saponins with important biological activity, enhanced stability and toxicity has also led to the exploration the structure-activity relationships. The great structural diversity of saponins due to their various biosynthetic origins, the challenges of identification, the novel biological activities, possibility of developing potent herbal drugs for human society, novel biosynthetic processes and semi and total synthesis of saponins are now opening up new opportunities or newer trends for exploitation of novel saponins. Several reviews on triterponoied saponins have been published in recent years [1-5]. This review is concerned with the latest developments in isolation and structure elucidation of triterpenoied saponins and contains a compilation of the 13C NMR data of the novel aglycones of triterpenoid saponins, published during mid-1996-March, 2007 and arranged skeleton wise, together with the biological activities reported.
The usual methods of isolation of triterpenoid saponins have been reviewed [6-8]. The recent developments in chromatographic techniques have been employed to isolate pure individual triterpenoid saponin successfully. As an example the bioactive saponins of the Indian medicinal plant Bacopa monniera were successfully isolated as follows. The air dried powdered whole plants were first defatted with petroleum ether (60-80°C) and then successively extracted with CHCl3 and MeOH. The methanolic extract was partitioned between n-BuOH and water. The n-BuOH soluble fraction of the methanol extract was partially purified by adsorption on silica gel followed by successive elution with CHCl3, EtOAc, acetone and 20% MeOH in CHCl3. The ethyl acetate and acetone extracts on chromatographic purification followed by preparative HPLC (μBonadapak C18 column) yielded five relatively non polar saponins. The CHCl3-MeOH extract was chromatographed on silica gel and then subjected to preparative HPLC to yield four polar saponins [9-10]. Patagonicoside A, a novel antifungal disulfated triterpene glycoside from the sea cucumber Psolus patagonianswas isolated as follows. The sea cucumbers, frozen prior to storage were homogenized in EtOH and centrifuged. The dried ethanolic extract was partioned between MeOH-H2O (90:10) and cyclohexane. The methanalic extract was subjected to vacuum dry column chromatography on Davisil C18 reversed phase column (35- 70 μ) using H2O and H2O -MeOH mixtures. The fractions thus obtained were further purified by reverse phase HPLC to give the pure saponin [11].
Structure elucidation
Structure elucidation of the isolated pure triterpenoid saponins is usually carried out by a combination of chemical and spectroscopic methods. The advent of modern spectroscopic methods, mainly 2D NMR and soft ionisation mass spectroscopy, allows rapid structural study of saponins and examination of small amount of the intact saponin prior to any treatment which might produce artifacts. The recent introduction of HPLC coupled to UV photodiode array detection (LC-DAD-UV), mass spectrometry (LC-MS or LC-MSMS) and NMR (LC-NMR), and of LC-coupled bioassay provide online useful structural information of plant constituents with only a minute amount of plant material avoiding the time consuming isolation of pure components [12]. Renukappa et al. have applied LC-NMR (liquid chromatography-nuclear magnetic resonance) and LC-mass and LC-coupled bioassay to determine two anthelmintic dammarane-type triterpenoidal saponins, significantly active against Caenorhabditis elegans from a crude fraction of Bacopa monniera [13]. Nyberg et al. also applied solid phase extraction followed by NMR and MALDI–TOF mass spectrometry on chromatographic fractions QH–A and QH– B of immuno adjuvant active saponins to identify 28 different saponins of Quillaja saponaria [14].
Mass spectroscopy
The molecular masses of triterpenoid saponins are determined by different soft-ionization mass spectrometric techniques such as FABMS, fields desorption, plasma desorption and laser desorption [15-19]. A liquid chromatography/mass spectrometric (MS) method involving electrospray ionization together with up front collision induced dissociation (CID), tandem MS or MSn fragmentation has recently been developed for rapid online characterization of triterpene saponins [20].
The molecular masses, the number of monosaccharide units and their sequence, and also the identification of key fragments related to the structural variation at C-3 and C-28 oligosaccharides of ten quillaja saponins were determined by nanoelectrospray ion trap multiple stage tandem mass spectrometry. In positive and negative mode ESI-ITMS mostly the [M+Na]+ ion at m/z 1591.8 and the ion [M-H]- at m/z 1567.9 were obtained for the bisdesmoside saponin 6. The former displayed the fragment ions at m/z 1121.8 [M+Na–GlcA-Gal-Xyl]+, 653.3 [Fuc+Rha+Xyl+Rha+Na+H2O]+, 635 [Fuc+Rha+Xyl+Rha+Na]+and 489.2 [Fuc+Rha+Xyl+Na]+, together with those given by glycosidic bond cleavage. The ion [M+ GlcA+Gal+Xyl-H]- at m/z 955.5 showed loss of the C-28 oligosaccharide. The ion at m/z 361.1 derived via retro Diels Alder fragmentation also appeared [21]. Liu et al. discussed the ESI-ITMS for structure elucidation of triterpene saponins in Polygala tenuifolia [22]. In order to study the fragmentation pathways of the triterpenoid saponins from Bacopa monniera. Zehl et al. applied different fragmentation techniques of MSI-ITMS, low energy CID, PSD and high energy CID. The fragment ions provided the sequence and branching of the aglycone part as well as the molecular mass of intact aglycone [23].
NMR spectrometry
Of all the physical methods, the NMR technique has changed most during the last three decades. With the introduction of fast multidimensional 2D and 3D NMR techniques and also computer assisted. Structure Elucidation programs the structural studies of saponins without derivatization, especially for the identification of monosaccharide units, determination of their anomeric configuration; their sequence and the position of glycosidation have been greatly facilitated. Consequently the structural information of saponins is more readily obtained than by any other single technique. High field NMR experiments viz COSY, HETCOR, COLOC and NOESY were used for the determination of the complete structure of the novel antifungal disulfated triterpene glycoside from the sea cucumber Psolus patagonicus [11]. The 13C NMR spectrum displayed 56 carbon signals. A DEPT experiment revealed 7-CH, 13–CH3, 12 –CH2, 7– quaternary carbon resonances. Correlation of 13C signals with those of directly bonded protons was achieved by means of 2D HETCOR experiments. The proton and carbon signals for the sugar units were assigned by means of COSY, HETCOR and COLOC. The NOESY spectrum of 1 displayed the correlation between H-12/ H-9, H-9/H-19, and H–12/H-21. As a result 12- OH and 17-OH were shown to be α while S configuration is at C-20. The sugar–sugar and sugar- aglycone linkages were also indicated by NOESY data (Figure 1).
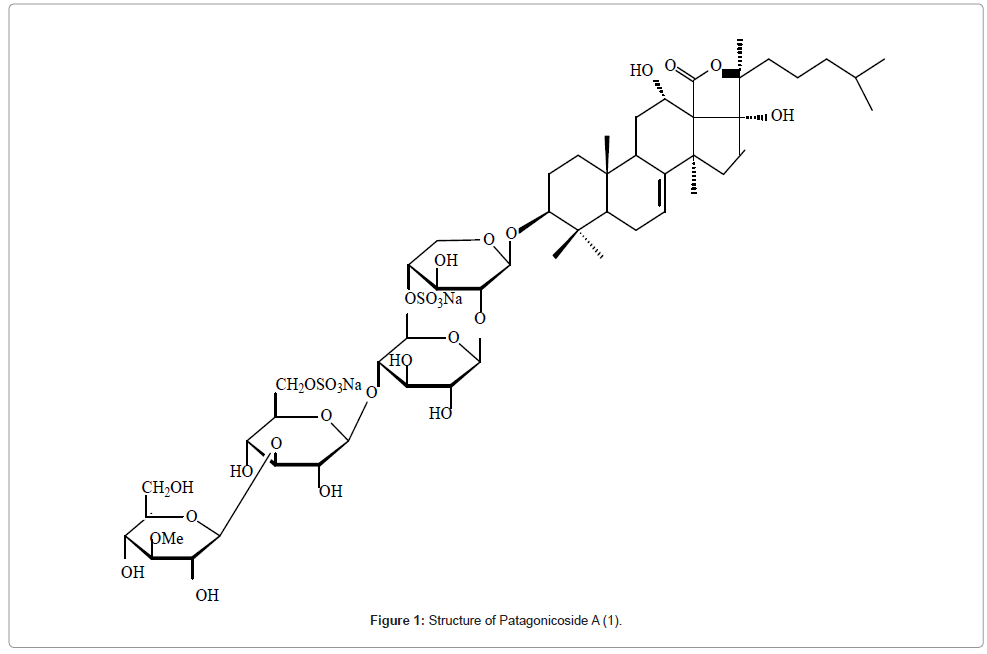
Figure 1:Structure of Patagonicoside A (1).
The structures of six triterpenoid saponins obtained from Maesa balansae were elucidated by the techniques such as COSY, NOESY, HMQC, HMBC, TOCSY and DEPT [24]. For example saponin 2 showed in its 13C NMR spectrum five anomeric carbon signals and its aglycone had an oleanane skeleton with an oxygen bridge between C-13 and C- 28. These data suggested that 2 is a triterpenoid pentasaccharide. The assignments were confirmed by long-range coupling in HMBC and by spatial interaction in NOESY. The spatial proximities observed between H-3 and H-23, H-3 and H-5, and H-16 and H-28 suggested β and α configurations at C-3 and C-16 respectively. The correlation between H-18 and H-22, and H-21 and H-29 allowed assignment of the (Z)–cinnamoyl group to C-22 and benzoyl to C-21. The nature of the monosaccharides and their sequence were determined by a combination of DQFCOSY, TOCSY, DEPT, HSQC and HMBC. Starting from the anomeric protons of each sugar unit, signals for all the hydrogens within each spin system were assigned by DQFCOSY and TOCSY. On the basis of the assigned protons, the 13C NMR resonances of each sugar unit were identified by HSQC and further confirmed by HMBC experiment (Figure 2).
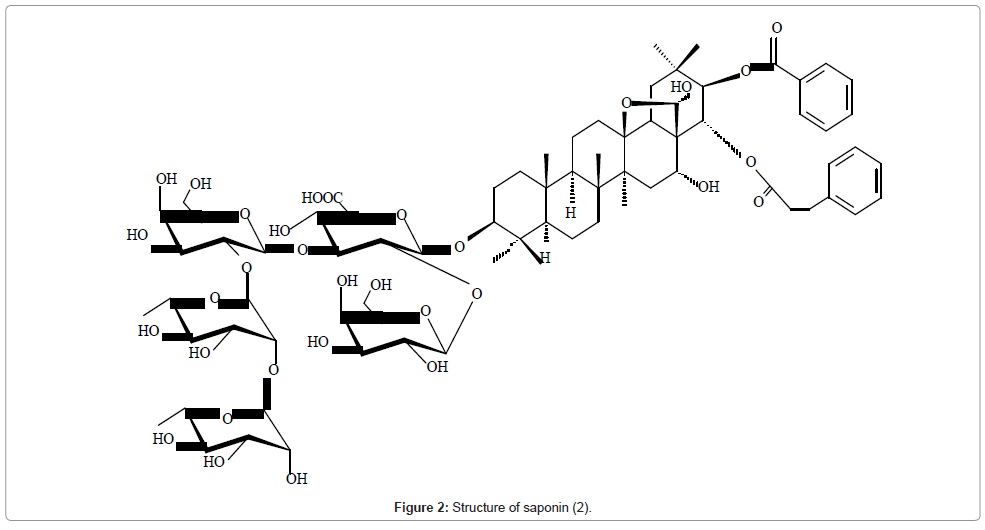
Figure 2: Structure of saponin (2).
The structures of two new dammarane–type triterpenoid saponins, bacopasaponins E and F isolated from the reputed Indian Medicinal Plant, Bacopa monniera were elucidated by spectroscopic methods and some chemical transformations [9]. The 13C signals of the two saponins were assigned and ring sizes of the sugars, mode of attachment and the sequence were determined by DEPT, 1H-1H COSY, HSQC and HMBC experiments (Figure 3).
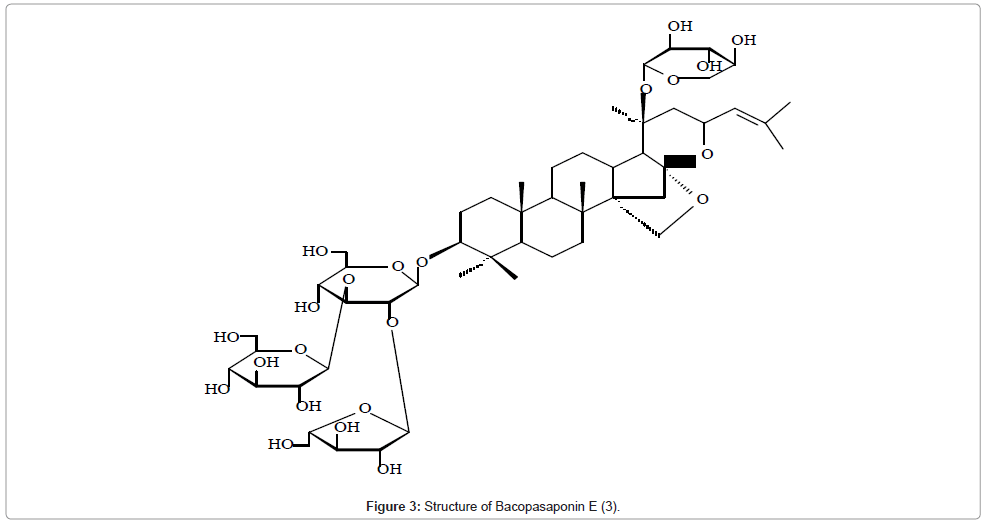
Figure 3: Structure of Bacopasaponin E (3).
The saponin polyacectyleneoginsinoside-Ro with an unknown profile of biological activity was isolated from Panax ginseng [25-30]. Its structure was established mainly by combination of 1D selective and 2D NMR techniques. The compound showed in its 13C NMR three anomeric carbon signals indicating the three sugar units. Charactesistic 13C values for one methyl carbon, seven methylene carbons, three oxy-methine carbons, two terminal double bond carbons, and two triple-bond carbons indicated a polyaccetylene group attached to a sugar unit. The configurations at C-3', C-9', and C-10' of the acetylenic part of the sugar unit were determined respectively as R, S, and S by comparison with the naturally occurring acetelenic compound panaxytriol using optical rotation and NMR data (Figure 4).
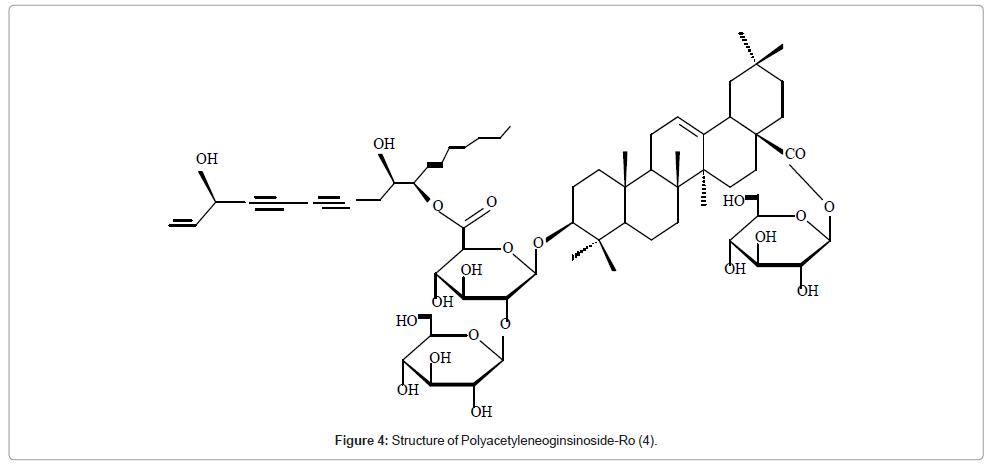
Figure 4: Structure of Polyacetyleneoginsinoside-Ro (4).
Reports of new triterpenoid saponins
A compilation of the 13C NMR data of the novel aglycones of new triterpenoid saponins arranged skeleton wise and reported during the period mid 1996-March 2007 are listed in Table 1. Structures 5-34 are aglycones of the various saponins and structure 35 is of a special saponin. New triterpenoid saponins isolated are listed in (Table 2 and Figure 5).
| Carbon No. | 23 | 32 | 31 | 34 | 18 | 24 | 22 | 16 | 17 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 39.0 | 39.0 | 38.9 | 32.4 | 39.7 | 39.3 | 39.8 | 38.8 | 38.5 |
| 2 | 26.9 | 26.5 | 26.7 | 30.2 | 26.6 | 25.0 | 26.6 | 26.5 | 26.4 |
| 3 | 88.5 | 89.6 | 88.8 | 89.1 | 82.3 | 80.7 | 91.3 | 88.9 | 88.4 |
| 4 | 39.5 | 39.7 | 39.5 | 42.6 | 44.0 | 42.7 | 39.8 | 39.6 | 39.4 |
| 5 | 55.8 | 55.8 | 55.7 | 54.1 | 48.2 | 48.5 | 56.6 | 55.9 | 55.8 |
| 6 | 18.5 | 18.4 | 18.4 | 68.1 | 18.9 | 18.5 | 19.3 | 18.6 | 18.3 |
| 7 | 38.4 | 34.9 | 34.8 | 38.5 | 33.5 | 35.3 | 34.8 | 33.2 | 35.7 |
| 8 | 40.9 | 41.8 | 41.7 | 46.9 | 40.8 | 42.4 | 41.2 | 40.0 | 41.9 |
| 9 | 51.4 | 50.3 | 50.6 | 21.3 | 52.0 | 49.5 | 48.1 | 49.7 | |
| 10 | 37.4 | 36.9 | 36.8 | 29.2 | 37.6 | 37.4 | 37.8 | 37.0 | 37.4 |
| 11 | 21.3 | 21.7 | 21.6 | 26.4 | 24.6 | 23.0 | 24.6 | 23.9 | 24.1 |
| 12 | 26.7 | 28.8 | 29.0 | 33.3 | 123.8 | 26.2 | 131.0 | 123.3 | 128.4 |
| 13 | 42.4 | 36.3 | 37.1 | 45.8 | 145.1 | 140.3 | 135.6 | 144.2 | 136.9 |
| 14 | 59.9 | 45.3 | 43.6 | 47.0 | 42.9 | 45.2 | 48.4 | 42.2 | 59.1 |
| 15 | 28.2 | 38.9 | 27.8 | 49.2 | 29.8 | 28.0 | 23.1 | 28.3 | 20.9 |
| 16 | 37.1 | 78.37 | 32.1 | 71.3 | 23.6 | 34.0 | 25.0 | 23.5 | 23.4 |
| 17 | 49.0 | 53.5 | 49.1 | 57.0 | 48.4 | 49.7 | 48.8 | 47.1 | 46.7 |
| 18 | 138.5 | 48.2 | 49.3 | 19.0 | 41.8 | 128.7 | 54.8 | 41.8 | 42.2 |
| 19 | 132.2 | 50.1 | 62.7 | 29.9 | 41.4 | 42.0 | 40.2 | 46.3 | 43.6 |
| 20 | 32.3 | 72.4 | 71.4 | 30.8 | 36.8 | 33.4 | 40.7 | 30.8 | 30.6 |
| 21 | 34.3 | 28.9 | 76.7 | 18.2 | 29.3 | 37.7 | 31.7 | 34.1 | 33.6 |
| 22 | 34.1 | 32.8 | 46.5 | 36.8 | 32.4 | 36.1 | 38.0 | 32.6 | 32.0 |
| 23 | 27.9 | 28.2 | 28.0 | 25.6 | 64.6 | 69.9 | 28.5 | 28.3 | 28.0 |
| 24 | 16.7 | 16.8 | 16.7 | 125.9 | 13.8 | 12.9 | 16.6 | 16.8 | 16.7 |
| 25 | 17.1 | 16.3 | 16.2 | 135.3 | 16.6 | 16.8 | 16.3 | 15.7 | 16.2 |
| 26 | 17.3 | 16.2 | 16.2 | 14.0 | 17.9 | 16.6 | 18.7 | 17.6 | 18.3 |
| 27 | 176.8 | 16.6 | 15.4 | 68.3 | 26.6 | 21.2 | 64.7 | 26.1 | 207.3 |
| 28 | 179.6 | 61.5 | 63.3 | 29.0 | 178.2 | 177.7 | 176.7 | 176.6 | 176.3 |
| 29 | 29.3 | 26.1 | 26.6 | 16.7 | 74.4 | 32.4 | 18.0 | 33.2 | 32.9 |
| 30 | 30.5 | 32.0 | 32.3 | 20.2 | 19.8 | 24.4 | 21.6 | 23.7 | 23.9 |
| [82] | [83] | [84] | [85] | [86] | [87] | [87] | [88] | [88] |
Table 1: 13C NMR data of aglycone part of the saponins.
| 35 | 28 | 29 | 30 | 19 | 14 | 15 | 12 | 13 | 20 |
|---|---|---|---|---|---|---|---|---|---|
| 31.5 | 47.5 | 39.7 | 39.9 | 39.3 | 38.0 | 37.3 | 45.7 | 45.5 | 38.3 |
| 25.4 | 66.5 | 26.9 | 26.9 | 28.0 | 24.6 | 24.6 | 36.3 | 36.3 | 26.7 |
| 79.0 | 95.3 | 89.6 | 89.6 | 75.7 | 83.0 | 83.4 | 79.6 | 79.5 | 85.1 |
| 38.4 | 41.0 | 39.8 | 39.8 | 54.7 | 54.4 | 54.4 | 44.1 | 44.1 | 53.4 |
| 42.6 | 56.5 | 56.8 | 56.9 | 52.2 | 48.2 | 47.9 | 52.2 | 52.1 | 51.7 |
| 19.1 | 18.6 | 18.5 | 18.6 | 21.9 | 19.9 | 19.7 | 20.6 | 20.6 | 21.4 |
| 39.6 | 34.8 | 35.7 | 35.8 | 33.2 | 32.0 | 31.7 | 35.5 | 35.5 | 32.8 |
| 45.7 | 40.8 | 40.9 | 40.9 | 40.5 | 38.4 | 39.3 | 42.3 | 42.3 | 41.5 |
| 51.5 | 55.0 | 51.3 | 51.3 | 48.6 | 46.7 | 46.2 | 57.1 | 54.0 | 54.8 |
| 36.6 | 38.3 | 37.1 | 37.3 | 37.1 | 35.8 | 35.6 | 148.8 | 147.7 | 36.4 |
| 37.7 | 40.1 | 21.8 | 21.9 | 24.1 | 23.3 | 22.9 | 67.9 | 77.7 | 127.1 |
| 208.1 | 210.9 | 25.8 | 26.4 | 123.0 | 117.0 | 122.1 | 127.9 | 122.8 | 125.9 |
| 147.8 | 56.0 | 43.4 | 45.1 | 144.6 | 142.1 | 142.1 | 145.4 | 149.0 | 137.1 |
| 143.2 | 56.0 | 50.7 | 50.3 | 42.3 | 42.3 | 47.0 | 43.5 | 43.1 | 42.4 |
| 134.2 | 32.1 | 31.7 | 31.9 | 28.5 | 43.5 | 46.3 | 27.8 | 27.8 | 37.0 |
| 130.4 | 24.8 | 27.4 | 28.1 | 23.6 | 212.9 | 212.0 | 22.3 | 22.6 | 25.3 |
| 131.7 | 42.6 | 46.0 | 45.5 | 47.7 | 49.1 | 46.4 | 37.0 | 36.9 | 48.7 |
| 17.0 | 15.9 | 15.7 | 15.8 | 41.4 | 36.4 | 44.2 | 43.1 | 43.2 | 132.1 |
| 23.4 | 17.5 | 16.6 | 16.7 | 41.2 | 42.3 | 46.1 | 46.1 | 46.2 | 40.8 |
| 139.1 | 81.4 | 79.1 | 81.2 | 36.6 | 30.0 | 30.4 | 31.0 | 31.0 | 32.7 |
| 12.8 | 22.3 | 179.4 | 178.4 | 29.1 | 37.1 | 34.1 | 34.4 | 34.3 | 37.1 |
| 83.0 | 40.4 | 40.7 | 39.2 | 39.2 | 22.8 | 20.7 | 32.1 | 31.8 | 32.0 |
| 33.5 | 24.0 | 74.2 | 75.3 | 181.0 | 209.4 | 209.0 | 27.4 | 27.4 | 180.2 |
| 120.5 | 125.9 | 125.5 | 124.1 | 12.5 | 10.3 | 10.5 | 16.0 | 16.1 | 12.1 |
| 135.4 | 130.8 | 138.5 | 139.6 | 16.4 | 15.0 | 14.9 | 110.1 | 109.6 | 18.8 |
| 26.8 | 25.8 | 25.6 | 25.7 | 17.7 | 16.2 | 17.0 | 16.4 | 16.5 | 16.7 |
| 19.0 | 17.9 | 18.2 | 18.3 | 26.3 | 24.9 | 26.4 | 25.9 | 25.7 | 19.9 |
| 28.8 | 28.2 | 27.8 | 28.0 | 176.9 | 76.4 | 76.6 | 175.5 | ||
| 22.5 | 17.5 | 16.8 | 16.9 | 73.9 | 32.7 | 32.8 | 33.3 | 33.2 | 32.2 |
| 25.2 | 17.7 | 16.6 | 16.4 | 19.9 | 24.1 | 22.9 | 23.7 | 23.8 | 24.3 |
| [89] | [90] | [91] | [91] | [92] | [93] | [93] | [94] | [94] | [95] |
Table 2: New Triterpenoid Saponins Isolated.
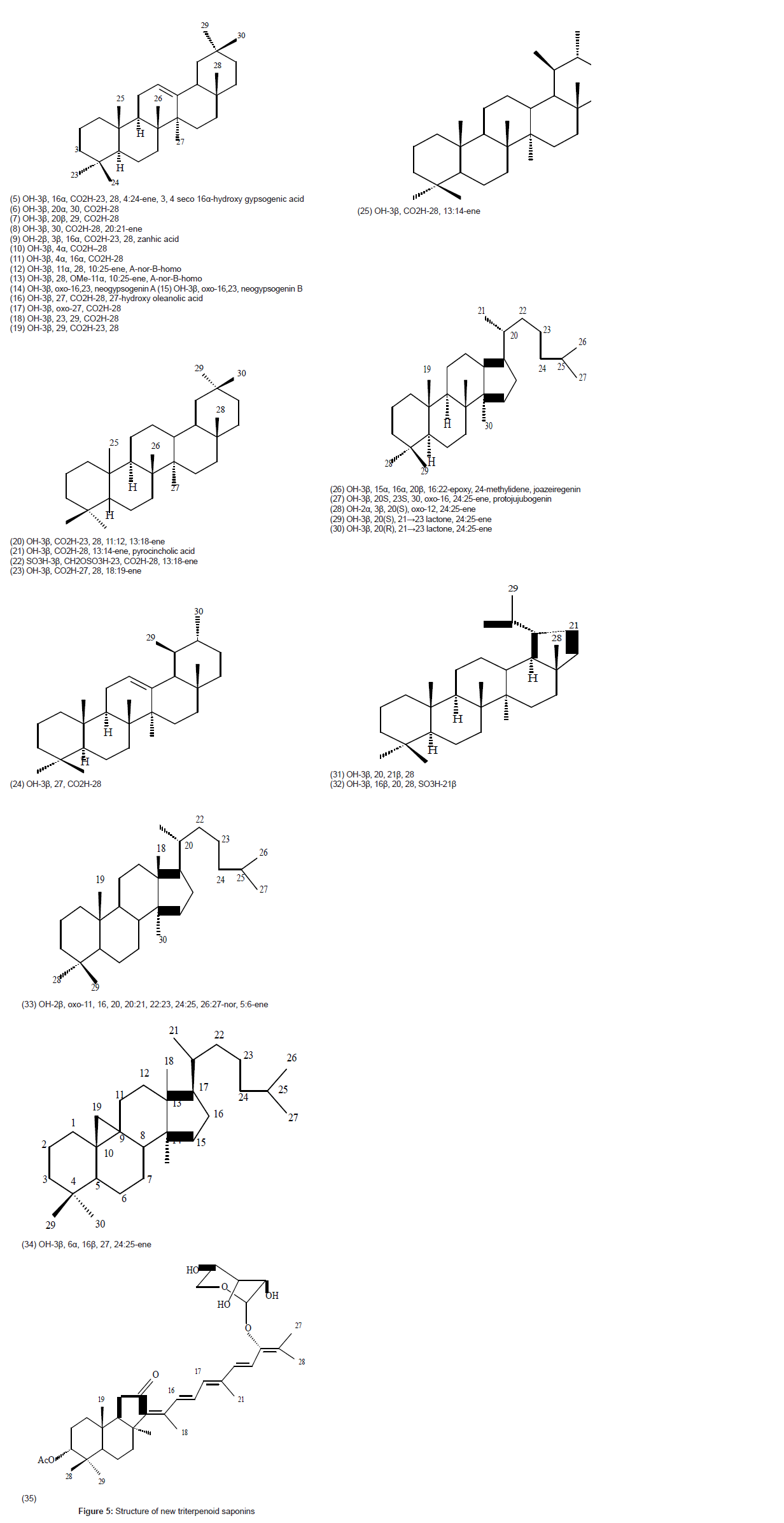
Figure 5: Representative photomicrograph of histopathological features in pulmonary necropsies.
Biological activity
Saponins have a wide range of biological activities that have attracted human interest from ancient times. Their toxicity is very low when administered to mammals, thus saponins present naturally in foods would be relatively non-toxic. Saponin containing foods could contribute to the cholesterol-lowering diets and increase the longevity of the individual’s lifespan. Vegetarians appear to be at least risk from cardiovascular disease. Earlier studies on the biological activities of saponins were limited to crude extracts containing saponins as well as other polar constituents. The advent of modern sophisticated methods of isolation and structure elucidation has attracted great interest of scientific community to study structureactivity relationships. A review on biological and pharmacological activities of triterpenoid saponins has been published earlier [31].
Antifungal activity
Saponins exhibits high toxicity against fungi. The antifungal properties of saponins are generally due to the ability to complex with sterols and cause membrane permeabilisation in fungi. Many triterpenoid saponins exhibit antifungal activity under experimental conditions. The triterpene glycoside, patagonicoside A of the sea cucumber Psolus patagonicus exhibited considerable antifungal activity against the pathogenic fungus Cladosporium cucumirinum[11]. The antifungal property of jujubogenin saponins of Colubrina retusa has been reported [32]. The triterpenoid glycosides, hyalodendrosides A and B isolated from a solid matrix fermentation of a lignicolous hyphomycete showed weak to moderate antifungal activity against some clinically relevant fungi [33]. Similar activity of triterpene glycosides, hermoiedemosides A and B from the Patagonian sea cucumber Hemoudema spectabillis was reported [34].
Molluscicidal activity
Schistosomiasis, commonly known as Bilharzia is a parasitic disease caused by thread worms of the genus Schistosoma with a mollusc as intermediate host. It is endemic throughout Africa, South America and several countries of Asia and affects millions of people in third world countries. The saponin containing plants responsible for molluscicidal activities are of great interest as they are inexpensive and ecofriendly compared to synthetic compounds. The hederagenin saponins of the soapnut, Sapindus mukorossi exhibited molluscicidal effects against golden apple snails Pomucea canaliculata [35]. The saponins desglucoanagalloside B and anagalloside B isolated from the plant Anagallis arvensis showed strong molluscicidal activity against Schistosome intermediate hosts, Biomphalaria glabrata and Oncomelania quadras [36]. The monodesmosidic saponins from Cestrum parqui possessed strong molluscicidal activity against Biomphalaria alexandrina snails [37]. The monodesmosidic saponin3-O- β-D-glucopyrananosyl (1→3)-α-L-rhamnopyranosyl (1→2)-β-D-xylopyranosyl oleanolic acid isolated from Dizygotheca elegantissimo has been found to be active against Biomphalaria alexandrina snails, the intermediate host of Schistosome mansoni in Egypt. Bisdesmosidic saponins had no snail kill activity [38]. The hederagenin saponins from the MeOH extract of Pometia eximia have been tested for molluscicidal activity. They revealed 100% mortality at 15 ppm of Biomphalaria glabrata and 84% mortality at 300 ppm of Aedes albopictuslarvae within 24 hour [39].
Antitumor/cytotoxic activity
Malignant gliblastoma is one of the most common malignant tumors in the neurological system. Cheng et al. investigated the proliferation change of human glioblastoma U87MG cells exposed to different concentrations (0.9-14.8 μM) of Tubeimoside V, a new cyclic bisdesmoside from tubers of Bolbostemma paniculatum. The compound significantly suppressed U87MG cell proliferation in a time and dose-dependent manner, with IC50 3.6 μM [40]. Kim et al. have isolated eight triterpenoid saponins from the root extract of Platycodon grandiflorum and evaluated the cytotoxic effect in vitro against human tumor cells like nor-small cell lung cancer, ovary cancer Sk-OV-3, melanoma SK-MEL-2, CNS cancer XF-498 and colon cancer HCT-15. The triterpenoid saponins exhibited significant inhibition on the proliferation of cultured cell [41]. Three new triterpenoid saponins, Julibrosides J29, J30 and J31 from Albizia julibrissin displayed significant anti-tumor activities in vitro against PC-3M-1E8, HeLa and MDA-MB-435 cancer cell lines at 10 μM assayed by SRB and MTT methods [42]. The novel triterpenoid saponin isolated from the root bark of Aralia dasyphylla has been found to show significant cytotoxic property against KB and Hela –S3 cells [43]. The novel acylated triterpene bisdesmosides isolated from the roots of Securidaca inappendiculata showed potent selective cytotoxic activity against M-CSF-stimulated macrophages and were suggested to have potential as new agents for the treatment of inflammatory diseases such as RA and atherosclerosis [44]. Mimaki et al. studied the cytotoxic activity of bisdesmosidic triterpene saponins from Enterolobium contortisiliquum against BAC1.2F5 mouse macrophages, EL-4 mouse lymphoma cells and L-929 mouse fibroblasts [45]. Contortisiliosides A and C were moderately cytotoxic to both BAC1.2F5 macrophages and EL-4 cells and contortisiliosides D-G did not show any apparent cytotoxic activities against the three cell lines. But contortisilioside B exhibited selective cytotoxic activity against BAC1.2F5 mouse macrophages with an IC3 value of 3.4 μM. The macrophage death caused by contortisilioside B was shown to be neither necrotic nor apoptosis-inducing. Three new triterpene glycosides intercedensides A, B and C from the sea cucumber Mensamania intercedens showed significant cytotoxicity against 10 human tumor cell lines with ED50 in the range 0.6-4.0 μg/mL. Intercedenside A exhibited significant in vivo antineoplastic activity against mouse Lewis lung cancer and mouse S180 sarcoma. It may be further studied as potential anticancer agents [46]. Enterlosaponin A isolated from Enterolobium contortisiliquum exhibited a highly selective cytotoxicity against BAC1.2F5 mouse macrophages and the macrophage death was shown to be neither necrotic nor due to induction of apoptosis [47].
The oleanane type triterpenoid monodesmosidic saponins were isolated from the n-BuOH extract of the roots of Clematis argentilucida and evaluated for cytotoxicity against human tumor HL-60, HepG-2, and SGC-7901 cell lines. They exhibited cytotoxic activity against the three cell lines with IC50 values in the range of 0.87- 19.48 μM [48]. Eight new triterpenoid saponins, Oleiferosides A-H isolated from the EtOH extract of the roots of Camellia oleifera were evaluted for cytotoxic activities against four human tumor cell lines (A549, B16, BEL-7402, and MCF-7) by using the MTT in vitro assay. Oleiferoside C exhibited potent cytotoxic activity against the tested cell lines with IC50 values <10 μM. Oleiferoside, A, B, D and E showed moderate cytotoxic activities towards the tested cell lines [49]. Li et al. [32] investigated the chemosensitizing activity of a diverse set of triterpenoid saponins in two human liver cancer cells (Bel-7402 and HepG2) and non-neoplastic HL-7702 liver cells. They identified cyclamin, a 13, 28-epoxy oleanane-type triterpenoid saponin isolated from Ardisia japonica as a potent chemosensitizer. The low cytotoxic level of cyclamin synergistically enhances the growth inhibitory effect of 5-fluorouracil (from 41.3 ± 1.1% to 64.5 ± 2.3%), Cisplatin (from 57.1 ± 50% to 79.6 ± 2.7%) and epirubicin (from 62.6 ± 1.2% to 74.9 ± 1.8%) on Bel-7402 cells, but not HL-7702 cells. Flow cytometric analysis showed that cyclamin also synergistically enhances the apoptosis and cell cycle arrest induction effects of 5-fluorouracil by 69.4% and 22.2% respectively. The mechanism of action could be through the activation of caspase 3, 8 and 9, the upregulation of cyclindependent kinase 2, the cell division cycle 25 homolog A expression level, or the Bax/B-cell lymphoma 2 ratio. Furthermore, a lactate dehydrogenase release assay demonostrated that cyclamin markedly increases the membrane permeability of Bel-7402 cells, which may also contribute to the mechanism of the chemosensitizing activity of cyclamin. These findings indicate the potential usefulness of cyclamin for the chemoprevention and treatment of liver cancer [50]. The effects of the oleanane-type triterpenoid saponin, DRβ–H isolated from Clematis ganpiniana, a Chinese traditional anti-tumor herb on growth and apoptosis of various human breast cancer cell lines was evaluated. DRβ-H showed strong inhibitory activity on the growth of various breast cancer cells and induced apoptosis in these cells. DRβ-H inhibited P13K/AKT and activated ERK signaling pathway. P13K inhibitor LY294002 synergistically enhanced DRβ-H induced apoptosis whereas MEK inhibitor U0126 reduced the apoptosis rate. Moreover, DRβ-H regulated the ratio of proapoptotic and antiapoptotic Bcl-2 family proteins. Furthermore, DRβ-H induced depolarization of mitochondrial membrane potential which released Apaf-1 and Cytochrome C from the inter membrane space into the cytosol, where they promoted caspase-9 and caspase-3 activation [51]. Han et al. isolated five oleanane type triterpenoid saponins glycoside ST-14a, glycoside ST-J, anhuienoside E, hedra saponin B and flaccidoside II from the rhizome of Anemone flaccida and evaluated for antitumor activity on human BEL-7402 and HepG2 hepatoma cell lines, as well as the model of cervical tumor cells (HeLa) treated with lipopolysaccharide (LPS). The triterpenoid saponins possessed potential antitumor activities and exerted their inhibition effects via inhibiting COX- 2/PGE 2 pathway [52].
Antidiabetic activity
The hypoglycemic effect of the saponin fraction of horse chestnut seed, Aesculus hippocastanum and of pure constituents escins Ia, Ib and IIa, on the elevation of plasma glucose level in the oral glucose tolerance test was examined in rats [53]. Escin IIa showed the most potent activity due to the presence of 2’-O-xylopyranosyl moiety in the oligosaccharide part and the acyl groups in the aglycone moiety. But betavulgarisides II and IV with the aglycone structure of oleanolic acid, isolated from the medicinal foodstuffs, sugar beet (root of Beta vulgaris) have been reported to exhibit hypoglycemic activity in rats at a concentration of 100 mg/kg in an oral glucose tolerance test [54]. The effect of the characterized triterpenoid saponins II and III from the rhizomes of Polygala senega var. latifolia in normal mice was studied by Kako et al. [55]. It was observed that the saponins II and III have dose dependent hypoglycemic activity. No change in blood glucose levels were observed in mice treated with the saponins IV. The triterponoid glycosides II-IV decreased the blood glucose level in NIDDM model with hyperinsulinemia. The hypoglycemic effects of II-IV were potent and glucose levels were similar to the basal level of normal mice indicating that those triterpene glycosides may affect insulin resistance of peripheral tissues. Two novel triterpenoid saponins from the roots of Aesculus assamica inhibited release of free fatty acids from epinephrine-treated rat adipocytes and enhanced glucose uptake into 3T3-L1 adipocytes [56]. Hu et al. [57] studied the anti-diabetic effects of triterpenoid saponins from Stauntonia chinensis on stimulating glucose uptake by insulin–resistant human HepG2 cells. Only the saponin 3-O–β–L–arabinopyranosyl(1→2) rhamnopyranosyl(1→3)- arabinopyranoside 30– norhederagenin significantly increased glucose uptake and glucose metabolism and also enhanced the phosphorylation of AMP-activated protein kinase (AMPK) and activated the insulin receptor (IR)/insulin recaptor substrate-(IRS-1)/phosphoinositide 3 kinase (PI3K)/Akt pathways. The triterpenoid saponins improved glucose up take and catabolism in hepatic cells by stimulating the AMPKand the IR/IRS–1/P13K/Akt signaling pathwaysand. The saponins can also enhance glucose uptake and insulin sensitivity, representing a promising treatment for type 2 diabetes mellitus [57]. The oleanane-type triterpenoid saponins (1 mg/mL) from the roots of Momordica cymbalaria increased (at concentration 1mg/mL) the insulin release in rat insulinoma cell line (RIN-5F) preexposed to adrenaline (5 μM) and nifedipine (50 μM). Pancreatic histology also indicated considerable quantitative increase in beta-cells (75%) when treated with the saponins. It was suggested that the oleanane-type saponins possessesed potential antidiabetic activity with respect to insulin secretion, which may be attributed to modulation of calcium channel and beta- cell rejuvenation [58]. Thao et al. investigated the effect of triterpenoid saponins isolated from the bioassay guided fractionation of the methanol extract of the root of Rosa rugosa Thunb on inhibition of sucrose related to diabetes mellitus (DM). The n-butanol fraction showed potent rat intestinal sucrose inhibitory activity with value of 87.62 ± 5.84% inhibition compared to the positive control acrabose (50.96 ± 2.97% inhibition at 0.02 mM). The triterpenoid saponins (1.0 mM) exhibited significant sucrose inhibitory activity, with inhibition percentage values of 41.17 ± 3.52, 46.80 ± 4.00, and 39.39 ± 4.19% respectively. The selected saponins also showed moderate sucrose inhibitory activity (ranging from 13.26 ± 7.00 to 32.08 ± 6.04% inhibition) at the same concentration. The data provides starting point for creating new sucrase inhibitors, which may be useful for the development of effective therapies for the treatment of DM [59].
Antileishmanial activity
Leishmaniasis is a parasitic disease caused by protozoa of the genus Leishmania, and is a major public health problem throughout most of the tropical and subtropical world. The triterpenoid saponins isolated from the methanolic extract of leaves of the Vietnamese medicinal plant Maesa balansae showed antileishmanial activity against visceral leishmaniasis (Leishmania infantrum amastigotes). This is much more active in vitro compared to the reference compound Pentostam which is currently used as first– line treatment for leishmaniasis [24]. Bacopasaponin C, an indigenous glycoside isolated from Indian medicinal plant Bacopa monaiera was tested for antileishmanial properties both in free state and in various delivery modes. It was found to be very active and without any side effects. Because of its indigenous origin and nontoxic nature, the compound could very well be considered for application (in India) in the clinic through these alternative delivery modes [60].
Miscellaneous effects
The hepatoprotective effects of three new oleanane glucuronides palustrosides I, II and III from Lathyrus palustris towards immunologically induced liver injury in primary cultured rat hepatocytes was examined. The hepatoprotective effects of palustrosides I and II were greater than that of palustroside III [61]. Bacopa monniera alcohol extract exerted a hepatoprotective effect against morphine induced liver toxicity [62]. The novel triterpene oligoglycosides from the roots of Bupleurum scorzonerifolium have been found to have hepatoprotective effect on liver injury induced by D–galactosamine (D-Gal N) and lipopolysaccheride (LPS) in mice [63]. Four new triterpenoid saponins isolated from the same source showed potent hepatoprotective effects on liver injury induced by the above agents and by D –Gul N in primary cultured rat hepatocytes [64]. The new triterpenoid saponins isolated from the leaves of Callicarpa nudiflora Hook were tested for hepatoprotective activities against D-galactosamine- induced toxicity and showed pronounced activity in WB-F-344 rat hepatic epithelial stem-like cells [65]. The isolated new triterpenoid saponins Schekwangsiensides A-G from Schefflera kwangsiensis exhibited moderate hepatoprotective activities against D-galactosamine induced HL-7702 cell damage [66]. The oleanane triterpenenoids from Terminalia arjuna, a cardioprotective drug exhibited a moderate free radical scavenging activity on hypochlorous acid production from human neutrophils [67]. The new triterpenoid saponins isolated from Ilex cornuta were tested against H2O2- induced H9c2 cardiomyocyte injury. They showed significant cell-protective effects. No significant DPPH radical scavenging activity was observed [68]. The cycloartane glycoside from the rhizome of Cimicifuga foetida showed anticomplement activity against the classical pathway of the complement system with an IC50 value of 29.6 μM. [69]. The adjuvant activity of six new oleanane– type triterpene bisdesmosides from Dolichos lablab has been reported [70]. Eight triterpenoid saponins from the seed of Aesculus chinensis were evaluated for their inhibitory activity against HIV-1 protease. They showed moderate inhibitory activity [71]. New uralsaponins M–Y have been isolated from the roots of Glycyrnhiza uralensis Fisch. The uralsaponins M, S, and T exhibited good inhibitory activities against the influenza virus A/WSN/33 (HINI) in MDCK cells with IC50 values of 48.0, 42.7, 39.6 and 49.1 μM, respectively, versus 45.6 μM of the positive control oseltamvir phosphate [72]. The ethanolic root extract of Codonopris lanceolata was evaluated for anti-inflammatory activity using the carrageenan induced rat hind paw edema model and displayed a significant activity of 51.82% inhibition at 200 mg/kg [73]. The new oleanane-type triterpenoid glycosides of Gymnema alternifolium showed antisweet activity [74]. Two new oleananetype triterpene glycosides of Helianthus annuus exhibited marked anti- inflammatory activity with the ID50 values in the range 65-262 mmol per ear [75]. The ginsenosides isolated from the root of Panax ginseng have been reported to exhibit most potent inhibitory activity on β-hexosaminidase release from RBL-2H3 cells and on the PCA reaction [76]. The synthesized ocotillol- type triterpenoid derivatives which are the main components of Panax ginseng were evaluated for their antibacterial activity. The two triterpenoid derivatives exhibited excellent antibacterial activity with MIC values of 1 μg / ml against Staphylococcus aureus and 8 μg /ml and 4 μg /ml against Bacillus subtilis, respectively. When the two triterpenoid derivatives were combined with two commercial antibiotics kanamycin and chloramphenicol they showed strong synergistic activity at sub-MIC levels against S. aureus USA300 and B. subtilis 168. Chloramphenicol turned from a bactesiostatic to a bacteriocidal agent when combined with one of the triterpenoid derivatives against B. subtilis 168 [77].
Biosynthesis of triterpene saponins and production of saponins by tissue culture
Plants synthesize a wealth of triterpene saponins as part of their growth and development. These are synthesized via the mevalonate pathway [78] followed by oxidation, substitution, rearrangement and glycosylations mediated by glycosyltransferases and other enzymes. Substantial progress has also been made in the identification and characterization of saponin glucosyl transfereses. A number of genes and enzymes involved in triterpenoid saponin synthesis have been discovered in Panax ginseng. Plant cell culture for large scale production of saponins economically in medicinally important plants has become more significant recently. Tissue culture of Gypsophila paniculata producing gypsogenin-3-O- glucuronide, commercially important as a good model to study the biosynthetic behaviour of triterpenoidal saponins, was established on different culture media with different concentrations of growth regulators to detect the differences in biosynthetic expression between callus and multiple shootline. The maximum saponin formation in shoot culture was observed when the medium contained 5 mg/1 IAA. The saponin content in callus tissue was highest when the medium contained 1.5 mg/1 each of 2,4 D and kinetin [79]. A basal liquid culture medium for saponin production in adventitious roots of panax hybrid (Panax ginserg and Panax quinquofolium) was examined. The root from seedling callui, cultivated in 1/8 Murashige–Skoog (MS) solid medium containing 2,4 dichloro pheroxy acetic acid was cultivated in Gamberg B5 liquid medium. The maximum saponin production of 2.87% was obtained in 1/8 MS medium for 8 weeks. The saponin contents were different in the cell culture and the cultivated plants [80]. The growth of ginseng and ginsenoside production in the presence of different genes of the TL-DNA and TR-DNA family of the Ri plasmid of Agrobacterium rhizogenesA4 in Panax ginseng hairy root cultures was studied. The highest biomass and ginsenoside production was obtained by some root phenotypes of Panax ginseng hairy root cultures [81].
Financial support from CSIR, New Delhi is gratefully acknowledged.
