Research Article - (2015) Volume 3, Issue 3
Leptadenia pyrotechnica has been commonly used traditionally for the treatment of tumors in Gulf countries. However, the molecular mechanism of the anti-cancer activity of Leptadenia pyrotechnica remains unclear. In the present study, 80% ethanolic extract of Leptadenia pyrotechnica and its fractions (n-hexane, EA, n-butanol and aqueous) were evaluated for in vitro cytotoxicity against colon cancer cell lines (HCT 116 wild type and HCT 116 p53−/−). The hexane fraction of Leptadenia pyrotechnica (LHE) is identified as the major anti-cancer fraction in Leptadenia pyrotechnica solvent extracts. Cells were incubated with different concentrations of LHE. Cell viability was quantified by crystal violet and MTT assays. Apoptotic cells were determined using Annexin V-propidium iodide (Annexin V/PI) staining and γH2AX assay for damaged DNA. Bax and Bcl-2, caspase-8, 9, 3 protein expression was analyzed by western blotting. LHE decreased cell viability in colon cancer cells in a dose- and time-dependent manner with an IC50 of 100 μg/ml after 24 h. Further experiments on colon cancer cells treated with LHE at 100 μg/ml for 24 h and 48 h revealed that it induces a p53-dependent apoptosis through intrinsic as well as extrinsic pathways. Analysis of DNA fragmentation by flow cytometry and H2AX assay showed a higher apoptotic cell death in p53 wildtype colon cancer cells treated with LHE in comparison to p53-/- cells. Data of western blots further demonstrated that LHE treatment caused activation of caspases, up regulation of Bax and down regulation of Bcl-2 in a time-dependent manner in HCT 116 cell lines. GC-MS analysis of LHE revealed the presence of derivatives of linoleic acid (octadecadienoic), diterpenes (phytol), triterpenes (squalene), and lupeol as the major constituents. These findings collectively suggest that LHE induces apoptosis in HCT 116 cells possibly mediated by ROS generation and involved multiple pathways including extrinsic, intrinsic and caspase-mediated pathways. Our results suggest that LHE may be applied as a potential anti-cancer drug against human colon cancer having intact p53 function.
Keywords: Leptadenia pyrotechnica; Apoptosis; p53; Oxidative stress; Caspase
Colorectal cancer is a major problem worldwide with the highest incidence in developed countries [1]. On a global scale, colorectal cancer is the fourth most common malignant neoplasm after lung, breast and prostate cancers [2]. Plants continue to be an invaluable resource of anticancer drug discovery in recent years. There has lately been an active move toward identifying biologically active substances with anti-cancer activity from traditional herbs [3].
Leptadenia pyrotechnica (Forssk.) Decne is a plant belonging to the family of Asclepiadaceae and is widespread in tropical Africa, Asia, the Mediterranean region and in the sandy plains in the Western Gulf countries. It is known in Arabic language as Markh [4]. Currently it is being cultivated in forests, farms and on the sides of roads [5]. It is commonly used traditionally for the treatment of a variety of inflammation-related diseases including rheumatism, asthma, wound healing and tumors [6]. The leaves and bark of the plant are used in folk medicine in Mali to prepare antispasmodic, anti-inflammatory and antibacterial remedies [7]. In United Arab Emirates, stem bark of Leptadenia pyrotechnica is also known to be used for the treatment of tumors [4]. Recently, it has been reported to have hypolipidemic and anti-atherosclerotic effects [8] and to be useful for the treatment of inflammatory bowel disease [9]. Previous chemical investigations of this plant have produced novel components such as pregnane glycosides and flavonoid glycosides [7,10]. In our previous study Leptadenia pyrotechnica and its sub-fractions revealed significant cytotoxic effect against MCF-7 human breast cancer cell line [11].
One obvious therapeutic goal in the treatment of cancer is to trigger apoptosis. The level of apoptosis in cancer cells has been considered a principal indicator of the anti-cancer activity of potentially therapeutic substances [2]. This genetically regulated form of cell death is a complex process that involves the active participation of affected cells in a self-destruction cascade and is defined by a set of characteristic morphological hallmarks, including membrane blebbing, shrinkage of cell and nuclear volume, chromatin condensation and nuclear DNA fragmentation [12].
In this context, the purpose of this study is focused on the chemical investigation of Leptadenia pyrotechnica major components and the study of anti-cancer effects of ethanolic extract of Leptadenia pyrotechnica and its sub-fractions against cultured human colon cancer cells. Further, we explored the anti-cancer mechanisms of Leptadenia hexane extract (LHE) on cultured human colon cancer cells for the first time.
Preparation of the plant extracts
A sample of the air-dried, ground aerial part of Leptadenia pyrotechnica (10 g) was extracted with 80% (v/v) aqueous ethanol (200 mL) and the mixture was macerated for 72 hours at 4°C. The resulting mixture was then filtered and divided into two equal portions. The first portion was dried under reduced pressure in a rotary evaporator at 40°C to give aqueous ethanol crude extract (6%). The other portion was concentrated under reduced pressure in a rotary evaporator and then suspended in water (50 mL). The mixture was extracted two times with n-hexane (2 × 100 mL) in a separatory funnel to give hexane fraction (LHE). The residue was further extracted with ethyl acetate (0.6%) and n-butanol (9%) along with the residual aqueous fraction (4%), respectively using a similar protocol. Each of the obtained fractions (hexane, ethyl acetate, n-butanol and water fraction) were dried, weighed, dissolved in DMSO (typically 25 mg/mL) and kept at -20°C for further analysis.
Analysis of LHE constituents by GC-MS
GC-MS analysis of LHE was carried out using Agilent 7890A GC with 5975C MS system equipped with HP-5MS 5% phenyl methyl silox fused capillary column (30 m × 250 μm × 0.25 μm). Oven program of 50°C for 5 min then 5ºC/min to 280°C for 15 min. Helium gas (99.999%) was used as a carrier gas at a flow rate of 1 mL/min. Injection volume of 0.5 μL was employed using a pulsed splitless technique. The injector temperature was maintained at 250°C and the ion-source temperature was maintained at 230°C. Oven temperature was programmed starting isothermally at 50°C for 5 min, with an increase of 5°C/min to 280°C (for 46 min) and ending with a 16 min isothermally at 280°C. The total GC-MS running time was 66 min. Solvent delay was 3.5 min. Mass spectra were recorded at 70 eV ionization capacity with a scan interval of 0.5 s and fragments from 40 to 570 Da. The software adopted to handle mass spectra and chromatograms was Agilent MSD ChemStation. Interpretation of mass spectra of all components was conducted using the database of National Institute of Standards and Technology library (NIST08.L). Accepted hits were considered if their spectra match with those of the library compounds at 90% or more. These hits were further verified manually for correctness. The relative percentage amount of each component was calculated by comparing its average peak to the total peak areas of all components.
Total phenolic contents and in vitro antioxidant capacity
The total phenolic as well as the total antioxidant capacity of Leptadenia pyrotechnica was determined by the methods of described in ref. [11]. The in vitro total antioxidant properties of LE were estimated by the ferric reducing antioxidant power (FRAP) and 2,2-azino-bis(3-ethylbenzothiazoline-6-sulfonate) (ABTS?+) assays. The FRAP assay is based on the reducing power of antioxidants in which a potential antioxidant will reduce the oxidized ferric ions to the ferrous ions. The ABTS assay is based on the reduction of blue-green (ABTS?+) by antioxidants to its original colorless ABTS form. In ABTS and FRAP assays, calibration curves of ascorbic acid were established, the antioxidant capacity of the LE was then expressed as μmol ascorbic acid equivalent/g dry extract.
Assaying of pro-oxidant activity in LHE
Pro-oxidants present in the LHE were determined according to the method described in ref. [13]. This method is based on the fact that oxidants present in the sample including hydroperoxides oxidize the ferrous ion–o-dianisidine complex to ferric iron. The resulting ferric ion makes a colored complex with xylenol orange in an acidic medium. The color intensity, which can be measured spectrophotometrically, is related to the total amount of oxidant molecules present in the sample. The assay is calibrated with hydrogen peroxide and the results are expressed in terms of μM hydrogen peroxide equivalent per liter (μMol H2O2 Equiv./L). Briefly, 50 μL of test sample was added to 1000 μL of reagent one (25 mM H2SO4, 150 μM xylenol orange, 140 mM NaCl and 1.35 M glycerol, pH was 1.8 ± 0.5), followed by adding 50 μL of reagent two (25 mM H2SO4, 5 mM ferrous ammonium sulfate and 140 mM NaCl and 10 mM o-dianisidine dihydrochloride, pH was 1.8 ± 0.5) and then incubated at room temperature for 3 min. The absorbance was read at 560 nm after the removal of flocculated material by centrifugation. The signal was read against a H2O2 standard curve and the results were expressed as μMol H2O2/L.
Culture of cells and maintenance
Wild-type and p53−/− HCT 116 (human colon cancer) cells were purchased from ATCC, Manassas, VA. Wild-type cells were cultured in RPMI1640, p53−/−cells in DMEM. Both media contained 10% fetal bovine serum, penicillin (100 U/ml), and streptomycin (100 μg/ml). The cells were maintained in a CO2 incubator at 37°C and 85 ± 5% relative humidity. These cells were sub-cultured and used for experiments.
Cell viability assays
As the cytotoxicity of many active ingredients of plants can be mediated by different toxic mechanisms, cell viability was measured here by the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazoliumbromide) method and crystal violet staining. Mitochondrial-dependent reduction of yellow MTT to purple formazan, as well as the intensity of crystal violet absorption both correlate to the number of living cells. In order to detect the cytotoxicity to HCT 116 cell lines (HCT wild type and HCT p53−/−), cells were plated and grown in 200 μl of growth medium for 20 hours in 96-well microtiter plates (7500 cells per well). These cells were then treated with ethanol extract of Leptadenia pyrotechnica (ET), Hex (hexane fraction), EA (ethyl acetate fraction), But (butanol fraction) and Wat (water fraction) at different concentrations (100, 200, 300, 400, 500, 1000 and 2000 μg/mL) and incubated for 24 and 48 h. The control group was treated with the equivalent amount of dimethyl sulfoxide (DMSO, the maximum of 0.2% of the assay mixture was used).
MTT assay is based on the formation of insoluble formazan by the reaction of MTT dye with active mitochondrial cells. The intensity of formazan dye was measured at 570 nm in a microplate reader; it is directly proportional to the number of living cells.
In the crystal violet assay, attached cells were stained with a 0.5% solution of crystal violet for 20 min. The dye was incorporated into attached cells; excessive crystal violet was washed away. After a drying period of 1 hour the bound dye was eluted from the cells with 95% methanol and read at 570 nm in 96-well microplate reader. Both methods yielded comparable results. Data are presented as means ± SD. All studies were performed in quadruplicates and repeated three times independently.
Possible role of p53 pathway in the action of LHE
In light of the important role of p53 in mitochondria-mediated apoptosis, it is possible that p53 may serve as a common target of LHE for initiating the apoptotic cascade. To directly test this possibility, we investigated the biological effects of LHE in a set of two HCT 116 cell lines: wild type and p53−/−.
Determination of apoptosis induction by Annexin V-PI staining
Apoptosis was measured using an Annexin V–FITC apoptosis detection kit (Roche Diagnostics, USA) which detected Annexin V bound to phosphatidylserine during the early stages of apoptosis, as well as propidium iodide (PI) staining for DNA. 1.5 × 106 cells/plate was grown for 20 h in 100 mm culture dishes. The cells were then treated with 100 μg/ml of LHE for 24 h and 48 h, control cells were treated with an equal amount of DMSO. The cells were then stained with the Annexin V–FITC kit according to the manufacturer's instruction. The distribution of viable (FITC/PI-negative), early apoptotic (FITCpositive), late apoptotic (FITC/PI-double positive), and necrotic (PIpositive, FITC-negative) cells were analyzed on a flow cytometer FACS Canto II (Becton-Dickinson, CA, USA) using a 488 nm excitation and 530/30 nm band pass filter for fluorescein detection and a long pass filter 2P670 nm for PI detection after electronic compensation.
Protein expression analysis
The HCT 116 cell lines (1.5 × 106/100 mm dish) were seeded, incubated overnight and then treated with LHE (100 μg/ml) for 24 h and 48 h. Cells were lysed in buffer containing 4M Urea, 0.5% SDS, 62.5 mM TRIS, 1 mM PMSF and 1% Protease Inhibitor Cocktail (Calbiochem), sonicated (30% amplitude, 12 pulses, 1 second each pulse) and then centrifuged (14000 rpm, 4°C, 10 min). The supernatant was used for the experiments. Protein content of the lysates was determined by Bio-Rad Dc Protein Assay (BioRad Laboratories, Hercules, CA, USA). The protein extract equivalent to 30 μg of whole protein of each sample was resolved on 12% SDS-PAGE at 30 mA/gel using Mini-PROTEAN® Tetra Cell electrophoresis system (Bio-Rad Laboratories, Hercules, CA, USA). Separated proteins were transferred onto 0.2 μm nitrocellulose membrane (Whatman, Germany) using Mini-PROTEAN® Tetra Cell electrophoresis system (Bio-Rad Laboratories, Hercules, CA, USA). The membranes were blocked for 1 h at room temperature in Tris buffered saline-tween-20 (1M Tris pH 7.4, 5M NaCl, 0.0005% tween-20) containing 5% dried skimmed milk powder. Membranes were probed overnight at 4°C temperature with the respective antibody (Bax, Bcl2, Caspase 3, Caspase 8, Caspase, PARP: Cell Signaling Technology Inc., MA, USA; γH2AX: Millipore Corporation, MA, USA; GAPDH-HRP: Abnova, Taipei City, Taiwan). These membranes were washed three times (5 min each) in TBST with gentle agitation, followed by incubation with HRP-conjugated secondary antibodies (ImmunoPure® goat-anti-mouse or anti-rabbit IgG peroxidase conjugated; Pierce, Rockford, IL, USA) for 1 h at room temperature. Protein bands were visualized using ImmobilonTM Western Chemilumenescent HRP Substrate (Millipore, MA, USA) according to the manufacturers protocol. Chemiluminescence image was captured using a Gene Gnome (Syngene, Cambridge, UK).
Results of the GC-MS analysis showed that at least 27 compounds were present in LHE (Table 1, Figure 1). Our results showed the presence of derivatives of linoleic acid (octadecadienoic, 6.19%), diterpenes (phytol, l5.17%), triterpenes (squalene, 8.75%, lupeol, 33%) in addition to some minor phytosterols in this fraction.
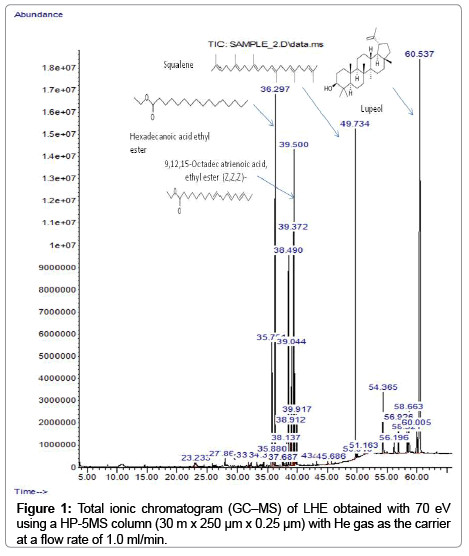
Figure 1: Total ionic chromatogram (GC–MS) of LHE obtained with 70 eV using a HP-5MS column (30 m x 250 μm x 0.25 μm) with He gas as the carrier at a flow rate of 1.0 ml/min.
| No. | Compound name | tR(min) | area (%) |
|---|---|---|---|
| 1 | 2-tetradecene, (E)- | 22.964 | 0.17 |
| 2 | 4,8-dimethyl-1,7-nonadiene | 23.237 | 0.59 |
| 3 | 1-hexadecene | 27.858 | 0.16 |
| 4 | tetradecanoic acid, ethyl ester(ethyl myristate) | 32.306 | 0.24 |
| 5 | 1,19-icosadiene | 33.223 | 0.15 |
| 6 | pentadecanoic acid, ethyl ester | 34.338 | 0.16 |
| 7 | n-hexadecanoic acid | 35.750 | 5.21 |
| 8 | ethyl 9-hexadecenoate | 35.874 | 0.69 |
| 9 | hexadecanoic acid, ethyl ester | 36.296 | 9.27 |
| 10 | heptadecanoic acid | 37.522 | 0.14 |
| 11 | 9,12-octadecadienoic acid, methyl ester, (E,E)- | 37.683 | 0.16 |
| 12 | heptadecanoic acid, ethyl ester | 38.142 | 0.57 |
| 13 | Phytol | 38.489 | 5.17 |
| 14 | 9,12-octadecadienoic acid (Z,Z)- (linoleic acid) | 38.910 | 1.92 |
| 15 | 9,12,15-octadecatrienoic acid, (Z,Z,Z)- (α- linolenic acid) |
39.046 | 5.97 |
| 16 | 9,12-octadecadienoic acid, ethyl ester (Z,Z)- (linoleic acid ethyl ester) |
39.368 | 6.19 |
| 17 | 9,12,15-octadecatrienoic acid, ethyl ester (Z,Z,Z)- |
39.504 | 6.21 |
| 18 | octadecanoic acid, ethyl ester | 39.913 | 1.21 |
| 19 | Squalene | 49.738 | 8.75 |
| 20 | lanostan-3-beta-ol | 51.163 | 0.13 |
| 21 | α -tocopherol | 54.360 | 2.51 |
| 22 | campesterol | 56.193 | 0.56 |
| 23 | stigmasterol | 56.924 | 1.69 |
| 24 | β-sitosterol | 58.324 | 1.29 |
| 25 | ismiarenol | 58.659 | 2.33 |
| 26 | hopenol B | 60.009 | 1.67 |
| 27 | lupeol | 60.542 | 33.21 |
Table 1: GC-MS results of the chemical constituents of LHE.
Total phenolic contents and in vitro antioxidant capacity
The in vitro antioxidant capacity of LHE was evaluated with FRAP and ABTS assays and compared with other anti-oxidant rich fractions of Leptadenia pyrotechnica reported in our previous study [11]. As shown in Table 2, the lowest level of total antioxidant capacity was found in the LHE fraction. The ascorbic acid equivalent antioxidant capacities of the LHE were 0.049 and 0.049 μmol/mg in FRAP and ABTS assays, respectively. Correspondingly, LHE also contained low amount of total phenolic content (10.53 mg gallic acid/g).
| Extract | Yield (%) | Total phenolic content (mg/g) | TAC, FRAP assay(µmol/mg) | TAC, ABTS assay(µmol/mg) |
|---|---|---|---|---|
| ET | 6% | 75.69 ± 4.39 | 0.235 ± 0.0005 | 0.449± 0.0005 |
| Hex | 2% | 10.53 ± 3.37 | 0.049 ± 0.0005 | 0.050 ±0.003 |
| EA | 0.6% | 252.27 ± 2.8 | 1.210 ± 0.020 | 0.568 ± 0.005 |
| But | 9% | 19.14 ± 0.94 | 0.080 ± 0.001 | 0.225 ± 0.001 |
| Wat | 4% | 17.76 ± 1.50 | 0.070 ± 0.001 | 0.185 ± 0.003 |
Values are means ± of three experiments. ET: Ethanol Extract; Hex: Hexane, EA: Ethyl Acetate, But: n-butanol and Wat: Water fraction.
Table 2: Total phenol content (gallic acid equivalent μmol/g dry extract) and total antioxidants capacity (TAC) (ascorbic acid equivalent μmol/mg) of ethanol extract and soluble fractions from Leptadenia pyrotechnica.
Assaying of pro-oxidant activity in LHE
In the pro-oxidant assay, ROS of LHE react with the ferrous ion– o-dianisidine complex to generate ferric ions, which can be detected by their ability to form complexes with xylenol orange in an acidic medium and generate a red chromogen. This ability to oxidize Fe2+ and stimulate the formation colored complex is adapted as a measure of the pro-oxidant properties of actual plant extract and the results were expressed as μMol H2O2/mg of extract. Figure 2 shows that LHE exhibited a pro-oxidant effect at the concentration used and increased the generation of ROS in concentration–dependent manner.
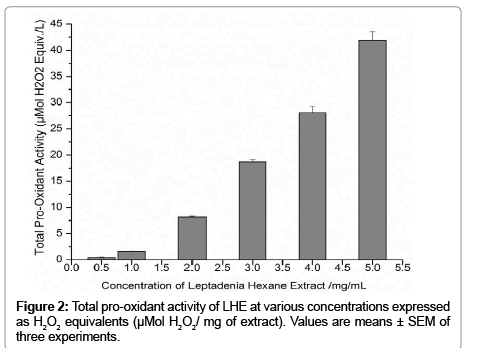
Figure 2 Total pro-oxidant activity of LHE at various concentrations expressed as H2O2 equivalents (μMol H2O2/ mg of extract). Values are means ± SEM of three experiments.
Effect of different fractions of Leptadenia pyrotechnica on cell viability in colon cancer cell lines
The cancer cell inhibition property of Leptadenia pyrotechnica extract was evaluated in wild-type HCT 116 human colon cancer cell line. In order to determine the effective concentration range, first we tested the dose dependence of Leptadenia pyrotechnica extract cytotoxicity measuring cell viability by MTT and crystal violet assays. Cells were treated for 24 h and 48 h with increasing concentrations (100–2000 μg/ml) of the crude ethanol extract of Leptadenia pyrotechnica (ET), Hex (hexane fraction), EA (ethyl acetate fraction), But (butanol fraction) and Wat (water fraction) and cell viability was measured. The Hex and EA extracts displayed better cytotoxic activity and 5B). Interestingly, the early apoptotic (Annexin V-positive and than the crude ET extract against HCT 116. Interestingly, Hex fraction exhibited the most pronounced effect. In fact, incubation of HCT 116 with Hex fraction for 24 h led to a dose-dependent cytotoxicity effect. Cytotoxicity of Hex fraction was most sensitively registered by crystal violet assay where the 50% lethal concentration (IC50) value after 24 h of incubation with Hex fraction was 100 μg/mL. The cytotoxicity of Hex fraction was lowest when tested with MTT while the same concentration (100 μg/mL) gave only about 30% cytotoxicity after 48 h and 500 μg/mL gave about 62% cytotoxicity after 24 h. Furthermore Hex fraction induced time dependent cytotoxicity of HCT 116 cells >90% loss of viable cells was observed after 48 h of treatment with 200 μg/mL in crystal violet assay in contrast to 500 μg/mL in MTT assay (Figures 3A and 3B). These findings demonstrate that different active ingredients in Leptadenia may give deferent results in MTT and crystal violet assays depending on their specific mechanisms of action.
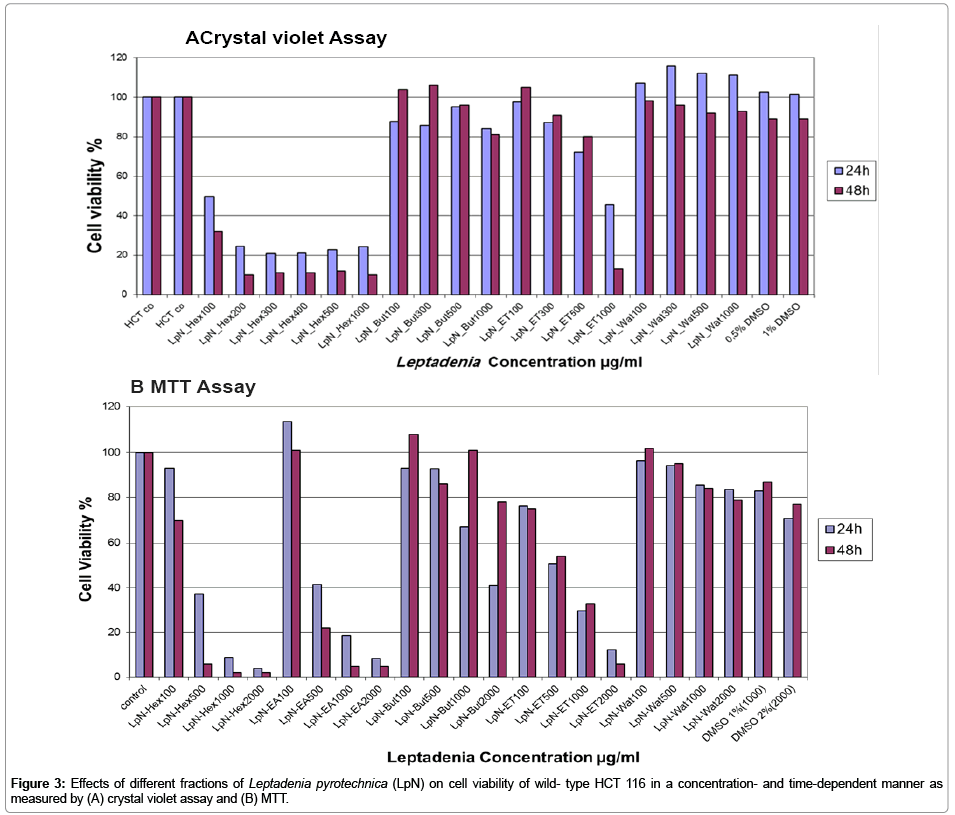
Figure 3: Effects of different fractions of Leptadenia pyrotechnica (LpN) on cell viability of wild- type HCT 116 in a concentration- and time-dependent manner as measured by (A) crystal violet assay and (B) MTT.
Possible role of p53 pathway and DNA double-strand breaks in the action of LHE
To elucidate possible mechanisms involved in the reduction in cell number by LHE we studied the expression of the DNA-damage marker γH2AX. Phosphorylated histone H2AX (γH2AX) appears during apoptosis and is an early chromatin modification following initiation of DNA fragmentation during apoptosis [14]. DNA doublestrand breaks were evaluated by the detection of γH2AX in wild-type HCT 116 treated for 24 h and 48 h with 100 μg/mL of Hex, EA and Wat fractions. The most remarkable increase in γH2AX expression in colon cancer cells was observed after 24 and 48 h of Hex treatment (Figure 4A).
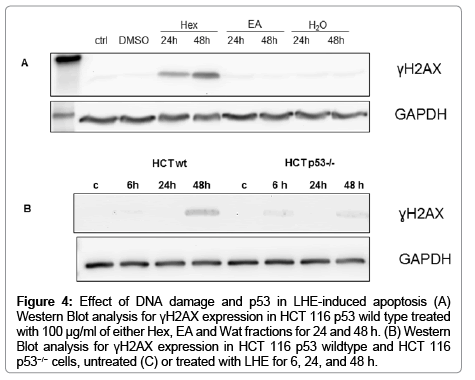
Figure 4: Effect of DNA damage and p53 in LHE-induced apoptosis (A) Western Blot analysis for γH2AX expression in HCT 116 p53 wild type treated with 100 μg/ml of either Hex, EA and Wat fractions for 24 and 48 h. (B) Western Blot analysis for γH2AX expression in HCT 116 p53 wildtype and HCT 116 p53−/− cells, untreated (C) or treated with LHE for 6, 24, and 48 h.
In light of the important role of p53 in mitochondria-mediated apoptosis, it is possible that p53 may serve as a common target of LHE for initiating the apoptotic cascade. To directly test this possibility, we investigated the biological effects of LHE in a set of two isogenic HCT 116 cell lines (p53 wild type and HCT 116 p53−/− cells). LHE significantly inhibited the proliferation of both colon cancer cells in a time- and concentration dependent manner. The most remarkable effects were observed at concentrations between 100 and 500 μg/mL. Compared to wild-type cells, cells deficient in p53 were resistant to growth arrest induced by LHE confirming the essential role of p53 in LHE-induced cytotoxicity (data not shown). In agreement with the result observed in Figures 4A and 4B shows that LHE at concentration 100 μg/mL induced extensive DNA fragmentation in p53 wild type cancer cells, but not in HCT 116 p53−/− cells. The close correlation between apoptosis and growth inhibition suggests that DNA damage might be required for LHE-induced expression of p53 as shown in Figure 4, and apoptosis is the key mechanism underlying the anti-proliferative action of LHE which is p53 dependent.
Pro-apoptotic effects of LHE
Cell death can be due to the non-specific unregulated process of necrosis or may be due to a programmed cellular death known as apoptosis which is being regulated by specific cellular signaling pathways. We investigated whether LHE causes a general non-specific necrotic cell death or induces apoptosis in both colon cancer cells (HCT 116) using Annexin V/PI staining technique. Figures 5A and 5B show that there was no increase in the necrotic cells after treatment with 100 μg/mL of HCT for 24 h. Treatment of cancer cells treated with LHE led to a change in the early stage apoptosis only in HCT 116 p53 wild type after 24 h of LHE treatment. After 48 h of treatment, a time-dependent increment in the number of apoptotic cells with LHE was observed in both cell lines compared to the basal apoptosis levels (Figures 5A and 5B). Interestingly, the early apoptotic (Annexin V-positive and PI-negative) and late apoptotic fraction (Annexin V-positive and PInegative) at 48 h was significantly higher in HCT 116 p53 wild type cells than in HCT 116 p53−/− cells (40% versus 10%, respectively), suggesting that the apoptotic effects of LHE is p53-dependent. We also characterized the apoptotic effect of both colon cancer cells by employing DNA double-strand breaks (Figure 4B).
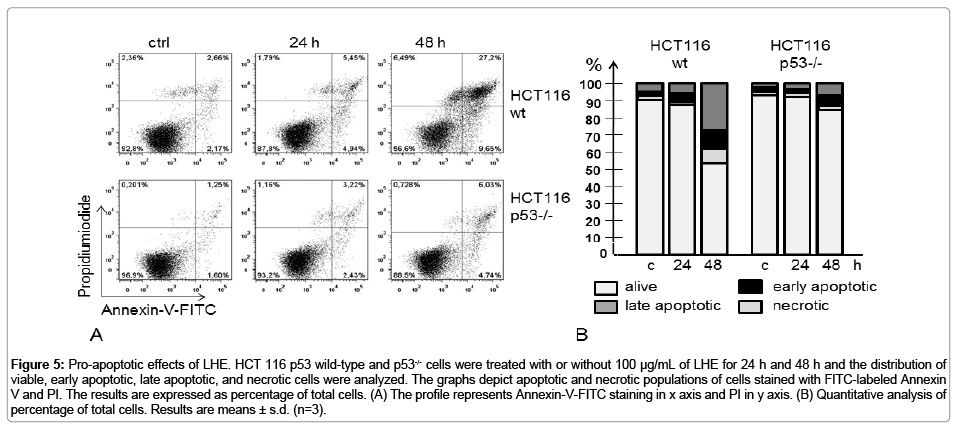
Figure 5: Pro-apoptotic effects of LHE. HCT 116 p53 wild-type and p53-/- cells were treated with or without 100 μg/mL of LHE for 24 h and 48 h and the distribution of viable, early apoptotic, late apoptotic, and necrotic cells were analyzed. The graphs depict apoptotic and necrotic populations of cells stained with FITC-labeled Annexin V and PI. The results are expressed as percentage of total cells. (A) The profile represents Annexin-V-FITC staining in x axis and PI in y axis. (B) Quantitative analysis of percentage of total cells. Results are means ± s.d. (n=3).
Effect of LHE on apoptosis related proteins expression of HCT116 p53 wild type and HCT116 p53−/−cells
In order to explore the potential mechanisms of apoptosis, expressions of major apoptotic proteins like Bcl-2, Bax, caspase 8, 9 and 3, and PARP were studied using western blot analysis. Results as depicted in Figure 6 showed clear evidence of apoptosis involvement in the induction of cytotoxicity by the extract. Bax was significantly higher in HCT 116 wild type cells at 48 h of LHE 100 μg/mL. As expected Bax protein expression was not detectable in p53−/− cells as Bax is known as one of the major transcriptional targets of p53. As shown in Figures 4B and 4C, LHE exposure of HCT 116 p53 wild type cells resulted in cleavage of caspase-3, -8 and -9. Caspase activation subsequently induced proteolytic cleavage of nuclear enzyme poly (ADP ribose) polymerase (PARP), which serves as biochemical marker of cells undergoing apoptosis. Interestingly HCT p53−/−cells did not show any PARP, caspase 9 and caspase 3 cleavage. The results indicate that LHE-induced apoptosis is caspase-mediated and p53-dependent. Interestingly, the more resistant HCT 116 p53−/−cells showed an upregulation of the anti-apoptotic protein Bcl2 under LHE exposure after 48 h. In contrast, in HCT 116 p53 wild type cells the Bcl2 protein was completely down regulated after 48 h.
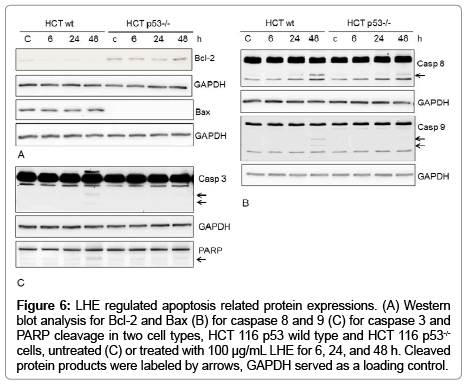
Figure 6: LHE regulated apoptosis related protein expressions. (A) Western blot analysis for Bcl-2 and Bax (B) for caspase 8 and 9 (C) for caspase 3 and PARP cleavage in two cell types, HCT 116 p53 wild type and HCT 116 p53-/- cells, untreated (C) or treated with 100 μg/mL LHE for 6, 24, and 48 h. Cleaved protein products were labeled by arrows, GAPDH served as a loading control.
In this study, we investigated the molecular mechanisms by which Leptadenia pyrotechnica hexane extract (LHE) induced anti-cancer effects and apoptosis in colon cancer cells. Our results provide the first evidence for cytotoxicity mediated by LHE through the induction of ROS accumulation, DNA damage, and caspase activation leading to HCT 116 colon cancer cell death in a p53-dependent manner. Interestingly, extracts of Leptadenia pyrotechnica, a perennial plant native to Gulf countries commonly used in traditional Emirates food and folk medicine, have been documented to have anticancer properties in human MCF-7 breast cancer cells [11]. In this study we explored the anti-cancer activity of the LHE on HCT 116 colon cancer cells having different p53 status. We demonstrate that out of five Leptadenia extracts; LHE was both the strongest inhibitor of cell proliferation and inducer of apoptosis in a dose and time-dependent manner. The level of apoptosis in cancer cells has been considered a principal indicator of the anti-cancer activity of potentially therapeutic substances [2]. Apoptosis is distinguished from necrosis by characteristic morphological changes, such as cell shrinkage, chromatin condensation, DNA fragmentation and exposure of phosphatidylserine on the outer layer of the plasma membrane. We used Annexin V-FITC and PI to discriminate early apoptotic cells from late apoptotic and necrotic ones, based on the translocation of phosphatidylserine to the outer layer of the plasma membrane of early apoptotic cells which is stained by Annexin V-FITC. We observed that LHE treatment of both colon cancer cells increased early and late apoptotic cell population after 48 h of treatment with IC50 concentration (100 μg/mL) with a significantly pronounced effect in HCT 116 wild type cells. In addition, LHE induced time-dependent DNA double-strand breaks presented by the detection of γH2AX in p53 wild type cancer cells. One of the earliest DNA damage responses is the formation of γH2AX as a result of double-strand breaks. γH2AX is formed in the chromatin before phosphatidylserine externalization is detectable in the outer plasma membrane of apoptotic cells [14]. This indicates that LHE-induced early DNA fragmentation and apoptosis is partly dependent on p53 activation.
In light of the important role of p53 in mitochondria-mediated apoptosis, the feature of p53 deficient HCT 116 cells prompted us to investigate the p53-independent mechanisms in LHE-induced apoptosis. The widely studied p53 downstream responses include cell cycle arrest, DNA repair and apoptosis [2]. The p53-mediated transactivation of apoptosis-related genes includes Bcl-2 family members and the apoptotic protease activating factor-1 (APAF-1) [2]. In addition to the mechanism involving p53-dependent transactivation of apoptotic genes, p53 can also directly bind to and inhibit the Bcl-xl, Bax, and Bcl2 proteins, leading to the release of cytochrome C and the initiation of caspase cascade [15].
LHE treatment resulted in an increase in pro-apoptotic Bax expression and a decrease in anti-apoptotic Bcl-2 expression in a time dependent manner, confirming the involvement of the mitochondrial pathway in LHE-induced apoptosis in HCT 116 p53 wild type cells. The pro-apoptotic Bax protein is directly responsible for making pores in mitochondria, but is blocked by the anti-apoptotic Bcl-2 homologues (Bcl-2, Bcl-xl, Mcl-1, A1, Bcl-w) [16] and [17]. A low expression level of Bcl-2 has been observed in cells undergoing apoptosis, and naturally occurring substances have been reported to induce apoptosis in carcinoma cell lines by decreasing Bcl-2 expression [18] and [12].
The activation of caspases is directly responsible for many of the molecular and structural changes in apoptosis [16,19]. Indeed, LHE treatment increased protein expressions of caspase-8 (initiator caspase for the death receptor pathway), casapase-9 (initiator caspase for the mitochondrial pathway) and caspase-3 (as major effector caspase for the two pathways) in time-dependent manner in HCT 116 p53 wild type cells (Figure 4). This indicates that LHE can cause both extrinsic and intrinsic apoptosis pathways. In contrast p53-/- cells did not show caspase 9 activation. Activation of caspases often leads to the proteolytic cleavage of several target proteins. These include degradation of DNA repair enzyme (PARP) and DNA dependent protein kinase, and cleavage of chromatin at inter-nucleosomal sites mediated by caspase-activated DNase [19,17]. Supporting it, LHE treatment led to a time-dependent increase in γH2AX expression (Figure 2) and PARP cleavage in HCT 116 p53 wild type cells (Figure 4C). This confirmed that caspases play a pivotal role in induction of apoptosis by LHE when p53 function is intact.
Induction of apoptotic DNA fragmentation could be induced by ROS which are determined here in the LHE as total pro-oxidant activity. Polyphenols have been reported to be able to exert enzymatic and/or chemical auto-oxidation and accelerate reactive oxidant species production [20]. Green tea polyphenols have been shown to inhibit viability of cancer cells by inducing apoptosis, an effect attributed to ROS (H2O2 and superoxide anion) formation [21]. In the present work, the pro-apoptotic effects of LHE may be attributed to a prooxidant activates of their polyphenols and induction of oxidative damage to tumor cell DNA. These results suggest that LHE-mediated pro-apoptotic effects are at least partially dependent on a pro-oxidant activates of the polyphenols in LHE.
Results from GC-MS analysis revealed that LHE contained unsaturated aliphatic compounds with lupeol being the main component (33%). This pentacyclic triterpenoid was previously found to have anti-proliferative effects against various cancer cell lines. Recent reports showed that lupeol directly inhibit tumor growth, cell cycle progression, and induce the apoptosis of tumor cells under in vitro and in vivo situations [22,23]. However, it is interesting to find that three chemical constituents in LHE can contribute either directly or indirectly to the treatment of cancer due to their potentiation effect. These identified compounds in our hexane fraction namely; squalene [24], hexadecanoic acid, ethyl ester, phytol [25] and a derivative of linoleic acid (octadecadienoic acid) have been reported to inhibit the growth of several tumor types [26-32].
In conclusion, this is the first study that demonstrates the biological mechanisms of LHE-induced apoptotic effects in colorectal HCT 116 cells. Apoptosis was more pronounced in tumor cells expressing p53. Our results also suggest that the early event involved in the apoptotic process might be associated with the generation of ROS and DNA damage. Moreover, LHE-induced apoptosis is attributed to extrinsic and intrinsic caspase-mediated pathways in a p53-dependent manner.
The authors are grateful to Maria Leidenberger for her excellent technical assistance.
